Первыми крупными шаровидными кактусами, с которыми познакомились европейцы при открытии Америки, были мелокактусы, в изобилии произрастающие на Антильских островах. Как правило они образуют одиночный стебель средних размеров от сдавленно-шаровидной до коротко-цилиндрической формы с хорошо выраженными широкими и высокими ребрами и крепкими шиловидными, прямыми или изогнутыми, колючками. Мелокактусы хорошо отличаются от других своих собратьев наличием очень своеобразного цветоноса - "цефалия" - генеративного побега на верхушке стебля. Цефалий не имеет хлорофилла и устьиц для газообмена в поверхностных тканях, очень густо окутан щетинками и опушением и предназначен исключительно для цветения и плодоношения. Даже неопытные любители, увидев однажды взрослый мелокактус, затем уже никогда не спутают его с другими кактусами.
Некоторую путаницу попытался совершить Курт Бакеберг (1960), объединив мелокактусы и дискокактусы в надродовой по своему иерархическому положению таксон "Sippe 10" подтрибы Boreocereinae Backbg. (1960). Однако ошибочность такой комбинации слишком очевидна, чтобы обсуждать ее еще раз, тем более, что это уже было сделано на страницах этого журнала в прошлом году (Филатов, 1999). В целом мелокактусы настолько однотипны, что не возникает сомнений в их монофилетичности, то есть в происхождении их от одного общего предка. В систематическом плане мелокактусы обособлены от других шаровидных кактусов. Ближайшими их родственниками являются цереусы: Coleocephalocereus и Buiningia.
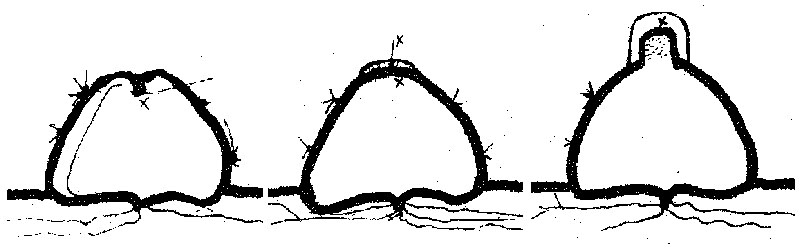
Интересно развитие цефалия. У молодых растений он отсутствует. Образование его начинается по достижении мелокактусом определенного возраста и размера. При этом, как мы предполагаем, апикальная меристема вегетативного побега расходится кольцом от центра меристемы, освобождая место апикальной меристеме цефалия. Одновременно изменяется и положение апикальной меристемы в стебле: "вегетативная" меристема находится ниже вдавленной макушки стебля и защищена от воздействия внешней среды тесно сдвинутыми основаниями ребер (рис. 2). Генеративная меристема, образуя большое количество щетинок и опушения, защищающего апекс от внешнего воздействия, поднимается вверх и по мере развития поднимается над вегетативным стеблем (рис. 3-4).
Внешне начальная фаза образования цефалия выглядит как появление в точке роста на верхушке растения небольшого количества щетинок и опущения, которые можно сперва даже не заметить, но сразу же вслед за появлением этих признаков начинается цветение растения. А это уже трудно не заметить, т.к. яркие дневные цветки мелокактусов, несмотря на небольшие в общем размеры, что называется, бросаются в глаза. Упомянутое появление первых признаков образования цефалия означает, что деятельность апикальной меристемы мелокактуса полностью перестроилась. Если на ювенильной стадии развития мелокактуса апикальная меристема образовывала "нормальный" вегетативный кактусный стебель с ребрами, то с вступлением в генеративную стадию развития мелокактуса меристема начинает образовывать верхушечный цветоносный побег - цефалий, совершенно непохожий на вегетативный.
В-четвертых, цефалий, в отличие от вегетативного стебля, не способен к фотосинтезу.
Очень большое количество ареол на поверхности цефалия обусловливает интенсивное и продолжительное цветение мелокактусов. По количеству цветков, измеряемых сотнями, которые образуют мелокактусы в течение хотя бы одного вегетативного периода (наблюдения проводились нами над культивируемыми растениями), им нет равных среди других шаровидных кактусов. Это понятно, поскольку количество новых ареол на цефалий, вырастающих за вегетационный сезон, в десятки, а возможно и в сотни раз превышает количество новых ареол на стеблях обычных кактусов.
Интересен вопрос, являются ли цефалии- цветоносы атавизмом, сохранившимся от "докактусных" предков - например, у толстянковых и портулаковых, родственных кактусам семейств, терминальные соцветия являются скорее правилом, чем исключением, или же цефалий являются неким новым органом, возникшим вследствие эволюцион- ного развития. Предварительные исследования показывают, что правильным является второе предположение. Более того, мы можем утверждать, что образование цефалия является прогрессивным направлением эволюционного развития, по той причине, что оно позволяет мелокактусам многократно увеличить семенную продуктивность по сравнению с "обычными" кактусами, имеющими похожие по форме и количеству семян плоды, например, по сравнению с маммилляриями.
Цветки мелокактусов сравнительно мелкие. Длина их варьирует в промежутке 15-43 мм, ширина - 4-25 мм, однако цветки абсолютного большинства видов не превышают 30 мм в длину и 16 мм в ширину. Большая часть длины цветка скрыта в глубине опушения цефалия. Цветки развиваются очень быстро и поэтому бутоны новых цветков можно заметить иногда всего за несколько часов, и не более, чем за сутки до начала цветения. Цветение отдельного цветка продолжается всего несколько часов в дневные часы, иногда до вечера. Венчики различных оттенков красного цвета - от розового до фиолетово-красного. Цветки самоопыляющиеся.
В природе возможно и перекрестное опыление, которое осуществляется главным образом колибри. Однако эффективность перекрестного опыления с помощью колибри снижается из-за очень небольшой площади кормового участка отдельных птиц. Это ведет к отсутствию генетического обмена даже между близко расположенными популяциями одного вида, что, в свою очередь, ведет к изоляции и появлению отчетливых различий между растениями разных популяций. Впрочем, несмотря на орнитофилию, цветки мелокактусов, как отмечали некоторые наблюдатели, посещаются также мелкими пчелами, которые более эффективно содействуют перекрестному опылению. Возможно, что в опылении цветков мелокактусов участвуют и другие насекомые. Так, Н. Тэйлор (1991) отметил посещение цветков Melocactus curvispinus в Венесуэле большой желтой бабочкой. Посещения бабочками мелокактусов были отмечены и в Восточной Бразилии, однако неясен эффект от этих посещении, так как возможно, что бабочек привлекает белый цефалий (Тэйлор, 1991).
Самофертильность цветков большинства видов мелокактусов способствует экспансии этих видов, так как даже изолированные одиночные экземпляры имеют возможность продуцировать полноценные семена. Самостерильность цветков отмечена в культуре только у М. azureus и М. glaucescens, что, возможно, объясняет причину очень ограниченного распространения этих, надо заметить, наиболее красивых видов рода, в природе. Два таксона мелокактусов имеют, подобно фрайлеям, клейстогамные цветки - М. lanssensianus и одна из форм М. peruvianus. Яркие красные плоды мелокактусов охотно поедаются в частности, птицами, которые, таким образом, содействуют распространению мелокактусов на большие расстояния.
Мелокактусы имеют очень большой ареал, уступая в этом отношении только опунциям. Ареал рода включает в себя юго-западную и юго-восточную прибрежные части Мексики, Центральноамериканские страны: Гватемалу, Гондурас и Коста-Рику, Антильские острова, Венесуэлу, Колумбию, Эквадор, Перу, Гайану, Суринам, Северную и Восточную Бразилию. Центром видового разнообразия рода является Восточная Бразилия, где на npocтранстве от Рио-де-Жанейро до широты г. Форталеза отмечены 15 из 31 видов рода, признаваемых в настоящее время (N.P.Taylor, 1991). Только в Восточной Бразилии можно найти участки местности, где совместно или вблизи друг от друга растут разные виды мелокактусов. На таких участках встречаются естественные гибриды. Также именно в Восточной Бразилии растут ближайшие родственники мелокактусов - колеоцефалоцереусы и буинингии. Все это вместе неопровержимо свидетельствует о том, что здесь и находится родина мелокактусов, откуда они распространились в другие регионы Америки.
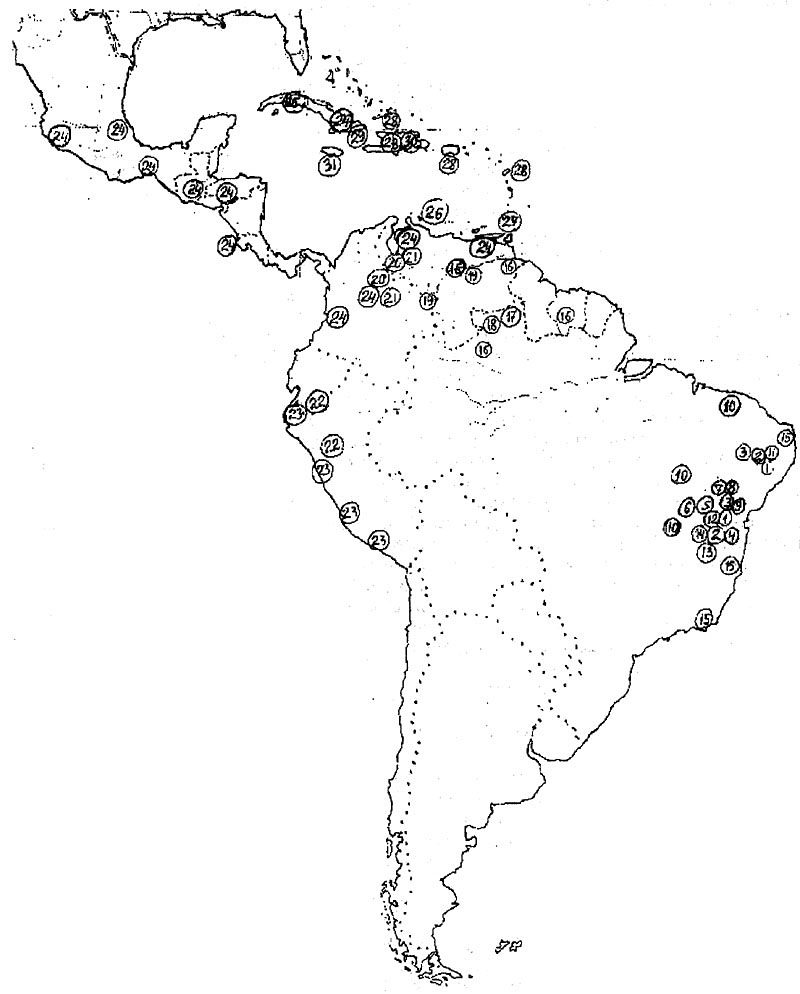
Рис. 5. Ареал рода Melocactus (по N.P.Taylor, 1991). Номера на карте соответствуют порядку описания видов в тексте.
В географическом распространении мелокактусов есть любопытный факт: четыре вида: Melocactus neryi, М. smithii, M. estevesii и M. mazelianus растут во влажнотропических районах северной Бразилии (штат Амазонас), Центральной и Южной Венесуэлы и Гайаны. В этих районах мелокактусы поселяются на открытых местах, а иногда и под пологом леса на обнажениях гранитных или гнейсовых пластов, на высотах до 650 м н.у.м. В большинстве этих районов наблюдается избыточное количество атмосферных осадков в течение всего года. В других, несмотря на сравнительно высокую годовую сумму осадков (1500- 3000 мм), отмечается выраженный сухой или засушливый сезон продолжительностью 1-4 месяца. На родине мелокактусов в Восточной Бразилии ситуация с выпадением осадков совершенно иная - здесь сухо большую часгь года, при избыточном увлажнении 2-5 месяцев. И лишь на юге ареала количество осадков возрастает и в районе к северу от Рио-де-Жанейро влажно в течение всего года.
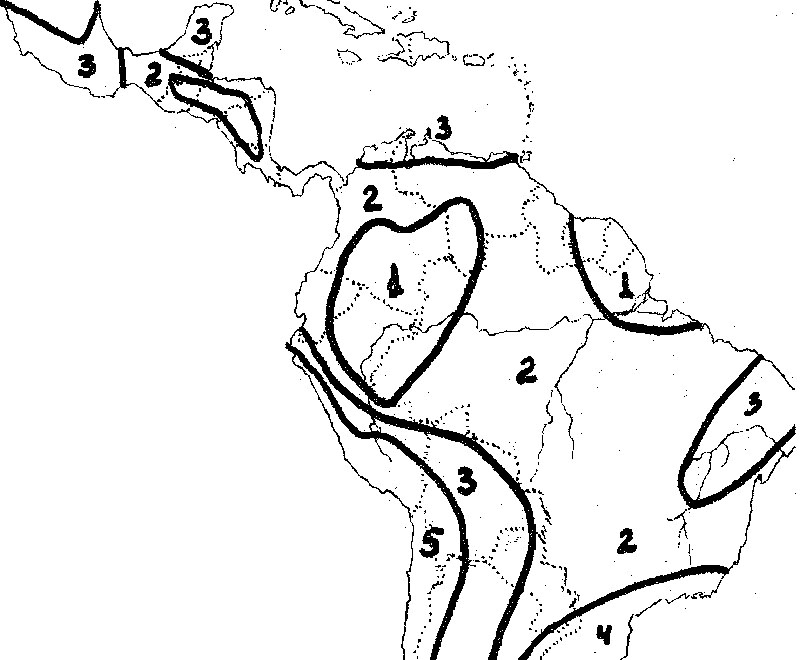
Рис. 6. Условия увлажнения в Центральной и Южной Америке (Агроклиматический Атлас Мира, 1972):
- Избыточное увлажнение в течение всего года.
- Избыточное увлажнение большей части года при наличии 2-5 сухих или засушливых месяцев.
- Сухо большую часть года при избыточном увлажнении 2-5 месяцев.
- Достаточное увлажнение в течение всего года.
- Сухо в течение всего года.
Говоря о количественном составе рода мелокактус, мы неизбежно сталкиваемся с проблемой вида, поскольку разное понимание смысла этого основополагающего понятия ботанической таксономии приводит к большой разнице в списке видов мелокактусов. Относительно мелокактусов эта проблема стоит достаточно остро с 70-х гг уходящего столетия, когда на страницы кактусных журналов хлынул буквально поток описаний новых видов мелокактусов. Большинство этих новинок было описано Буинингом и его коллегами, в первую очередь Бредероо. Очень интересно отметить при этом, что в одной из своих статей (прошу извинить, что не смогу дать ее библиографические данные) А. Буининг откровенно писал, что предпочитает сделать тщательное описание новинки, нежели проводить общепринятую в таких случаях кропотливую работу по её классификации, что предполагает, в частности, тщательное изучение гербарного материала уже известных видов. Естественно, что подобная позиция не могла не вызвать возражений со стороны профессиональных ботаников. Также естественно, что большинство новинок, описанных А. Буинингом и другими любителями и коммерческими сборщиками в качестве самостоятельных видов, не были признаны другими специалистами. В частности, неоднократно упомянутый выше Найджел Тэйлор из ботанического сада Кью (1991) признает самостоятельными лишь 8 видов из описанных А. Буинингом и Бредероо. Впрочем, этот ботаник хорошо известен своей "укрупнительной" позицией как в отношении видов, так и в отношении родов. Поэтому можно предположить, что в конечном итоге большая работа, проделанная в 70-х и 80-х гг "искателями кактусов" значительно больше увеличит видовой состав рода мелокактусов, нежели это признается в настоящее время Н. Тэйлором.
С мелокактусами связано одно очень устойчивое заблуждение, которое вот уже на протяжении по меньшей мере полутора столетий переходит из одного издания (например, Fцrster C.F., Rьmpler Т., 1886, ) в другое. Как повторяет его Д. Д. Мозет (Mauseth, 1989): "Необходимо подчеркнуть, что мелокактусы являются моноподиальными и цефалий является непосредственным продолжением ювенильного побега; переход от одного к другому, раз случившись, уже не может вернуться в прежнее состояние. Растения могут стать очень старыми, но цефалий будет расти, в то время как размеры ювенильной части стебля не меняются".
Автор этих строк не имел возможности изучать биологию мелокактусов в природе, но в свое время имел большую коллекцию мелокактусов, содержавшихся в условиях, приближенным к естественным - в сухом субтропическом климате Юго-Западного Туркменистана. Из примерно двадцати пяти видов мелокактусов, имевшихся в коллекции, 6 видов, каждый из которых имелся в коллекции в количестве не менее 4 экземпляров, образовали цефалий и цвели: М. amoenus, M. bahiensis, М. тахоnii, М. matanzanus, М. guitartii, M. peruvianus.
Наблюдения над мелокактусами в коллекции показали, что и после образования цефалия вегетативные стебли всех экземпляров мелокактусов продолжали расти. При этом довольно часто из-под цефалия образовывались новые ребра. Разумеется, это явление можно было бы отнести за счет влияния условий культивирования. Однако внимательное изучение фотографий мелокактусов в природных условиях позволяет предположить, что это же явление имеет место и в природе (например, Тауlоr, 1991: стр.35, 51, 53, 55, 68), особенно в тех случаях, когда цефалий формируется на ювенильных побегах, далеко не достигших присущих данному виду размеров. Мы можем дать следующее объяснение этому феномену: при определенных условиях апикальная меристема ювенильного побега не исчезает и не перестает функционировать после появления апикальной меристемы генеративного побега. Защищенная от воздействий внешней среды густым опушением нарождающегося цефалия, старая меристема формируется в виде кольца у основания этого цефалия и продолжает наращивать вегетативный побег из тканей цефалия. К сожалению, в силу не зависящих от нас обстоятельств наша коллекция после ликвидации Пархайского ботанического сада суккулентов в 1995 году была утрачена, наблюдения над мелокактусами прерваны. Поэтому мы не знаем, как долго может функционировать такая "кольцевая" меристема, если наше предположение соответствует действительности.
СИСТЕМАТИКА РОДА ПО Н. ТЭЙЛОРУ (1991)
Н. Тэйлор признает 31 вид мелокактусов. Все виды он подразделяет на 6 неформальных групп, т.к. не имеет необходимых оснований для выделения в составе рода формальных таксонов типа подрод, секция, подсекция, серия и подсерия. Для того, чтобы читатели могли использовать приводимую систему также в отношении "непризнаваемых" Тэйлором таксонов, которых значительно больше "признаваемых", мы приводим в настоящем изложении всю синонимику видов рода.
Группа Мelocactus oreas
1. Melocactus oreas Miquel (1840). Стебли от сдавленно шаровидных до удлиненных, 8-15 см высоты и 10-18 см в диаметре (здесь и далее первой указывается высота/длина, затем диаметр. Одиночное число без указания его характеристики означает высоту/длину), ребер 10-16 до 4 см ширины у основания стебля. Колючки иногда до 8 см. Цефалий до 12 х 4-8 см.
Синонимы: M.rubrisaetosus, M.oreas, M.oreas subsp. rubrisaetosus, M.oreas var. submunitis. В составе вида автор выделяет два подвида: Melocactus oreas subsp. oreas (Бразилия, штат Байя) и Melocactus oreas subsp. cremnophilus (Бразилия, штат Пернамбуко).
2. Melocactus ernestii Vaupel (1920). Стебли 9-45 x 7-22, ребер 9-13 до 7 см шириной у основания стебля. Центральные нижние колючки до 9, радиальные до 15 см длины. Цефалий 1 К х 8 см.
Синонимы: M.oreas subsp. ernestii, M.erythracanthus, M.oreas f.erythracanthus, M.azulensis, M.oreas f.azulensis, M.longispinus, M.oreas var. longispinus, M.interpositus, M.nitidus.
В составе вида автор выделяет два подвида: Melocactus ernesii subsp. ernestii (Бразилия, Пернамбуко) и Melocactus ernesii subsp. longicarpus (Бразилия, юг штата Байя, до северных р-нов Минас Жераис).
Синонимы подвида longicarpus: M.longicarpus, M.deinacanthus subsp.longicarpus, M.florschuetzianum, M.deinacanthus subsp. florschuetzianum, M.mulequensis, M.deinacanthus f. mulequensis, M.montanus, M.neomontanus.
3. Melocactus bahiensis (Britten & Rose) Lutzelburg. (1923). Стебель шаровидный, пирамидальный или сдавленно-шаровидный, 10-21 х 11-21, ребер 8-14, очень низких, до 6 см ширины у основания стебля. Колючек 9-16, как правило прямых.
Автор разделяет вид на 2 подвида: subsp. bahiensis и subsp. amethystinus. В составе первого выделены три формы.
Синонимы subsp. bahiensis: M.acispinosus, M.krainzianus, M.brederooianus, M.inconcinnus.
Синонимы subsp. amethystinus: M.amethystinus, M.lensselinkianus, M.griseoloviridis, M.glauxianus, M.ammotrophus.
4. Melocactus conoideus Buin. & Brederoo (1973). Стебель сдавленно-шаровидной формы, 10х17 см, ребер 11-14, очень низкие и округлые. Цефалий 4 х 7.5 см Бразилия, Байя.
Группа Мelocactus deinacanthus
Включает единственный вид, редкий в природе и эндемичный для открытых гранитных и гнейсовых склонов на юге штата Байя к востоку от р. Сан-Франсиску.
5. Melocactus deinacanthus Buin. & Вгеderoоо (1973). Стебель шаровидный до продолговатого, 15-35 х 12-25 см, ребер 10-12 до 4 см высоты и 6 см ширины у основания стебля. Ареолы крупные, до 10х15 мм. Колючек 15-21. Цефалий до 25х9.

Melocactus deinacanthus (5) фото N. Taylor.
Группа Мelocactus levitestatus
Включает один изолированный вид, эндемичный для выхода силурийских известняков на границе штатов Байя и Минас Жераис.
6. Melocactus levitestatus Buin. & Brederoo (1973). Стебель от шаровидного до цилиндрического, 15-68 х 14-30 см. Ребер 9-12,5х8 см. Эпидермис от светлого серо-зеленого до темно-зеленого цвета.
Синонимы: M.diersianus, M.securituberculatus, M.warasii, M.rubrispinus, M.diersianus f. rubrispinus, M.uebelmannii.

Melocactus lewitestatus (6) фото N. Taylor.
Группа Мelocactus azureus
Включает два вида очень красивых мелокактусов, имеющих чрезвычайно узкие ареалы.
7. Melocactus azureus Buin. & Brederoo (1971). Стебель от полуцилиндрического до цилиндрического, 13-45 х 14-19 см. Эпидермис от серо-зеленого до интенсивного голубого цвета. Ребер до 10, 35х40 мм. Колючек 9-15, черные или темно-коричневые до 5см. Цефалий 12х7-9см.
Синонимы: М.krainzianus, М.azureus var. krainzianus, М.ferreophilus.
Мelocactus azureus может быть подразделен на два подвида: М.azureus subsp. azureus и М.azureus subsp. ferreophilus.

Melocactus azureus (7) фото N. Taylor.
8. Melocactus pachyacanthus Buin. & Brederoo (1975). Стебель от сдавленно-шаровидного до овального 15-30 х 20 см. Эпидермис от светлого серо-зеленого до интенсивного голубого цвета. Ребер 9- 11, 25-37 х 50-65 мм. Колючек 10-12, все очень крепкие, красновато-коричневые. Цефалий 12 х 10 см.
Вид может быть подразделен на два подвида: M.pachyacanthus subsp. pachyacanthus и M.pachyacanthus subsp. viridis.

Melocactus pachyacanthus (8) фото N. Taylor.
Группа Мelocactus violaceus
Включает 9 видов мелокактусов, отличающихся острыми ребрами и небольшим количеством колючек в ареолах.
9. Melocactus salvadorensis Werdermann (1934). Стебель от пирамидального до сдавленно- шаровидного, 12-20х 12-25. Эпидермис сероватый, голубовато-зеленый, или интенсивный голубой. Ребер 8-14, Зх 5см. Колючек 8-14, толстых и коротких. Цефалий 15х6-1,0 см. Южные и восточные районы штата Байя в Бразилии.
10. Melocactus zehntneri (Britton & Rose) Lutzelburg (1923). Стебель от полушаровидного до цилиндрического, очень вариабельный, 11-48 х 13-25 см. Эпидермис зеленых оттенков, но иногда голубоватый. Ребра острые, колючек 8-13. Цефалий 11 х 6-10 см. Байя, Бразилия.
Синонимы: M.macrodiscus, M.сиrvicornis, M.zehntneri var. curvicornis, M.giganteus, M.helvolilanatus, M.canescens, M.canescens var.montealtoi, M.zehntneri subsp. canescens, M.zehntneri var.viridis, M.zehntneri var. ananas, M.arcuatispinus, M.douradaensis, M.saxicola.

Melocactus zehtneri (10) фото N. Taylor.
11. Melocactus lanssensianus P.J. Braun (1986). По-видимому, главным аргументом в пользу придания этому растению статуса самостоятельного вида являются клейстогамные цветки, которые самоопыляются не раскрываясь.
12. Melocactus glaucescens Buin. & Brederoo (1972). Стебель сдавленно-шаровидный или пирамидальный, в молодости светло-голубой, в старшем возрасте серо-зеленый, 13-18 х 14-24. Ребер 8-15,4 х 6 см. Колючек 6-10, цефалий до 10 х 7 см.
Синонимы: M.pruinosus и M.pruinosus sensu.

Melocactus glaucescens (12) фото N. Taylor.
13. Melocactus concinnus Buin. & Brederoo (1972). Стебель сдавленно-шаровидный, в молодом возрасте голубоватый, 8-12х 11-16 см, ребер 8-12, колючек 7-9, сперва красных, по мере старения чернеющих. Цефалий размерами 5 х 4-9 см. Растет на каменистых почвах в каатинге в штате Байя, Бразилия. Характерной чертой этого вида является зеленая окраска водозапасаюшей паренхимы внутри стебля.
Синонимы: M.axiniphorus, M.robustispinus, M.zehntneri subsp. robustispinus, M.pruinosus var. concinnus, M.macrodiscus.

Melocactus concinnus (13) фото N. Taylor.
14. Melocactus paucispinus G. Heimen & R. Paul (1983). Стебель дисковидной формы, 7-11 х 15- 19 см. Ребер 9-10, 4х5 см. Колючек 4-6, все радиальные. Цефалий 3-6 х 7-8 см. Байя, Бразилия. Молодые растения этого вида можно спутать с дискокактусами.
15. Melocactus violaceus Pfeiffer (1835). Стебель часто мелкий, темно-зеленый, никогда не бывает голубоватым, обычно диаметр больше высоты (5-18 х 6-17). Ребер 8-15, колючек 5-10. Цефалий 5х4-8 см.
Синонимы: Cactus melocactoides, M.melocactoides, M.depressus, M.melocactoides f.capensis, M.melocactoides f.exsanguis, M.melocactoides f.perspinosus, M.melocactoides f.sanctaritae, M.melocactoides f.depressuss, M.melocactoides var.itaipuassuensis, M.melocactoides var.natalensis, M.melocactoides var.violaceus.
Вид подразделяется на 3 подвида: M.violaceus subsp.violaceus, M.violaceus subsp.ritteri и M.violaceus subsp.margaritaceus.
Синонимы M. v. subsp. margaritaceus: M.margaritaceus, M.margaritaceus var. disciformis, M.margaritaceus var. salvadoranus, M.pentacentrus, M.depressus.

Melocactus violaceus (15) фото N. Taylor.
16. Melocactus neryi K. Schum. (1901). Стебель как правило сдавленно-шаровидный, 6-8 х 7-20 см, эпидермис темный, голубовато-зеленый. Ареолы глубоко утоплены в бороздках на острых ребрах. Колючек 7-11 Цефалий 5 х 4-9 см. Распространение: Зап. и Центральная Венесуэла, Северная Бразилия (Рио Арака и Амазонас) и Суринам.
Синонимы: Cactus neryi, M.guaricensis, M.schulzianus, Cactus caesius, M.amoenus.
17. Melocactus smithii (Alexander) Buining ex G. Rowley. Стебель чаще цилиндрический, нежели шаровидный, до 20 х 13-21 см, зеленый до желтовато-зеленого. Ребер 12-16. Колючек 9-10. Цефалий до 18х9 см. Произрастает в Сев. Бразилии (Рорайма) и в прилегающих юго-западных районах Гайаны на выходах гранитных пород.
Синонимы: Cactus smithii, Echinocactus amazonicus, M.roraimensis.

Melocactus smithii (17) фото N. Taylor.
Группа Мelocactus curvispinus
Включает 7 видов мелокактусов, отличающихся, в частности, особенностями тесты семян. Ареал группы находится за пределами Восточной Бразилии.
18. Melocactus estevesii P.J. Braun (1989). Стебель цилиндрический, 22 х 10-17 см, зеленый. Ребер 12-16, высоких и узких, 4 х 3 см. Колючек 9-14. Цефалий 13х6 см. Точное местообитание неизвестно. Вероятно, на скалистых обнажениях в северной Рорайме - Северная Бразилия.
19. Melocactus mazelianus Riha (1981). Стебель шаровидный, 9-27 х 7-20 см, темно-или желто-зеленый. Ребер 9-15. Колючек 6-14. Цефалий 12х8 см. Вост. Колумбия, Западная и Центральная Венесуэла.

Melocactus mazelianus (19) фото N. Taylor.
20. Melocactus schatzlii Till & Gruber (1982). Стебель шаровидный, 8-18 х 10-20, слегка голубоватый или серо-зеленый. Ребер 10-15, обычно острых. Колючки слабые, 8-16. Цефалий до 12 х 10 см. Сухие склоны долин и скалы в Андах на северо-востоке Колумбии и в прилегающих районах Венесуэлы на высотах 500-1050 м н.у.м.

Melocactus schantzlii (20) фото N. Taylor.
21. Melocactus andinus R. Gruber ex N.P. Taylor (1991). Стебель шаровидный 8-25 х 10-25 см, от темно- до желто-зеленого или желтый. Ребер 10-15 Колючек 8-17. Цефалий до 15 х 8 см. Местообитание: пологие, скалистые сухие склоны долин на высотах 1500-2200 (до 3000) м над у.м. в Андах в Колумбии и прилегающих районах Венесуэлы. Это самый "высокогорный" вид мелокактусов из ныне известных.
Синоним: М.gruberi.

Melocactus andinus (21) фото N. Taylor.
22. Melocactus bellavistensis Rauh & Backeb (1956). Стебель шаровидный, до 8-25 х 10-25 см., блестящий темно-зеленый. Ребер 9-18. Колючек 6-15 до 25 мм. Цефалий до 15 х 10 см. Н. П. Тэйлор уверен, что этот вид является более южным "родственником" двух предыдущих видов.
Melocactus bellavistensis может быть разделен на два подвида:
Melocactus curvispinus subsp. dawsonii.
Синоним: M.dawsonii. Распространен в штате Ялиско на юго- западе Мексики на прибрежных скалах. Эта популяция - наиболее далеко продвинутая на северо-запад. Подвид находится под угрозой уничтожения, т.к. имеет очень ограниченный ареал, в пределах которого быстро исчезает в связи с туристическим строительсгвом.
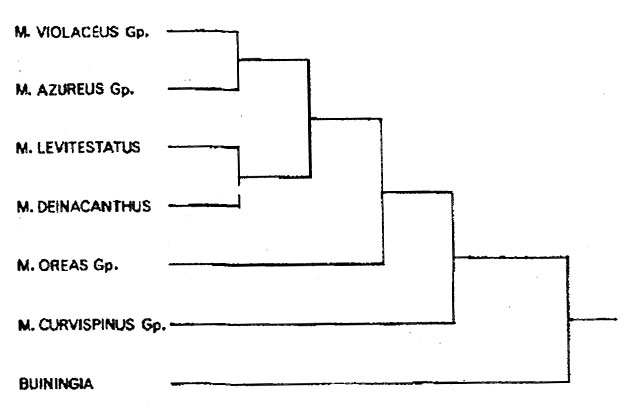
Дендрограмма родственных связей некоторых мелокактусов и буйнингии.
Виды островов Карибского моря, не вошедшие в группы
Семь видов не были классифицированы H. Тэйлором из-за отсутствия необходимого гербарного материала.
25. Melocactus matanzanus Leon (1934).
Синоним: М.actinacanthus.
Этот карликовый вид очень трудно отличить от венесуэльской формы Мelocactus neryi. Melocactus matanzanus немного меньше по размерам. Это один из наиболее популярным мелокактусов в коллекциях. Он сравнительно быстро образует красивый красный цефалий. В природе растет не на гранитах, а на известняках.
26. Melocactus macracanthos (Salm-Dyck) Link & Otio (1827). Очень вариабельный вид, распространенный на островах Кюрасао, Бонайре, Аруба и прилежащих мелких островах Нидерландской Вест-Индии.
Синонимы: M.barbarensis, M.bozsingianus, M.citrispinus.
27. Melocactus broadwayi (Br. & R.) A. Berger (1926). Распространен на островах Тобаго, Гренада, Гренадины и Сент-Винсент. Возможно, является промежуточным между Melocactus macracanthos и Мelocactus intortus.
28. Melocactus intortus (Miller) Urban (1919). Это наиболее крупный вид мелокактусов с наибольшим числом ребер - до 27, обычно 14-17. Распространен на островах Вест-Индии от Южных Багамских островов (Лонгайленд) до Мартиники. Вид полиморфен, что неизбежно при наличии большого количества изолированных популяций.
Синонимы: Cactus intortus, Cactus coronatus, Cactus communis, M.communis, M.amoenus, Cactus antonii, M.antonii, M.intortus var.antonii.

Melocactus intortus (28) фото Adams, "Bradleya", 7/89.
29. Melocactus harlowii (Br. & R.) Vaupel (1912). Вид из юго-восточной Кубы - провинции Ориенте.
Синонимы: Cactus harlowii, M.acunae subsp.lagunaensis, M.borhidii, M.evae, M.nagyi, M.radoczii.
30. Melocactus lemairei (Monville ex Lemaire) Miquel ex Lemaire (1840).
Синоним: Echinocactus lemairei.
Эндемик о. Гаити. Отличается очень мощными колючками, щетинками на цефалии, которые похожи на колючки, а также, иногда, полеганием своих цилиндрических стеблей.
31. Melocactus caroli-linnaei N. Р. Таylor (1991). Эндемичный ямайский вид.
Синонимы: Cactus melocactus, M.melocactus, M.communis, M.coronatus.
КУЛЬТИВИРОВАНИЕ МЕЛОКАКТУСОВ
Почти во всех книгах по кактусоводству содержится предостережение "начинающим" воздержаться от выращивания мелокактусов. Это предостережение вызвано тем, что все мелокактусы происходят из тропических и даже экваториальных районов, и поэтому они плохо переносят стандартные условия зимовки, применяемые, например, для мексиканских или аргентинских кактусов.
В природных условиях мелокактусы "зимуют", то-есть пережидают сухой сезон, при повышенных температурах и при повышенных уровнях солнечной радиации. Это противоположно тому, как зимуют в наших коллекциях "Мексиканцы". Поэтому первое правило при выращивании мелокактусов, это в период зимней стагнации содержать их при комнатной температуре. Холодный подоконник для мелокактусов совершенно непригоден. Конечно, у некоторых любителей они содержатся зимой именно на холодном подоконнике, вместе с другими кактусами. Но эти же любители потом признаются, что двадцатилетние мелокактусы у них все еще не образовали цефалия. Если в глубине комнаты места для мелокактусов нет, следует разместить их на полке в верхней части оконного проема (там намного теплее, чем на подоконнике), и так, чтобы рядом не было форточки.
Второе условие для успешной зимовки мелокактусов - это искусственное досвечивание растений 10-12 часов в сутки, причем желательно лампой с близким к дневному спектром, например такими, которые применяются для освещения улиц и подземных переходов.
И третье условие - не поливать.
Летнее содержание в целом несложно. Досвечивание не требуется. Растения надо обязательно разместить в теплице или тепличке, где дневная температура воздуха будет не ниже 30oC, а ночная - не ниже 20oC.
Необходимо предусмотреть отопление и летом на случай продолжительной холодной и пасмурной погоды, когда даже днем температура в вашей теплице не достигает нужного уровня. Если летом удастся обеспечить требуемые дневные (и ночные !) температуры, то никаких проблем с выращиванием мелокактусов у вас не будет.
Это быстрорастущие кактусы, очень устойчивые (как эхинопсисы) к переливу и постоянной влажности почвы. Мелкие виды, как, например, Melocactus niatanzanus, могут начать образование цефалия уже через 4 года после посева. Почва стандартная кактусная.
ЛИТЕРАТУРА