ЖУРНАЛ "КАКТУСЫ И ДРУГИЕ СУХОЛЮБИВЫЕ РАСТЕНИЯ" - выпуск 3 (2000 год)
ОСНОВНЫЕ БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ СУККУЛЕНТНЫХ ЭПИФИТНЫХ ОРХИДЕЙ
И.Е.Синев, Москва
Изучение суккулентных растений, таких как кактусы, стапелии, молочаи, толстянки, агавы, алоэ и другие, показывает, что все они характеризуются тремя основными признаками, которые в совокупно присущи только суккулентам:
- морфологически - наличием специализированных водозапасающих клеток в органах ассимиляции
- физиологически - САМ-типом фотосинтеза;
- экологически - засухоустойчивостью и способностью обитать в условиях периодически полн" отсутствия влаги в окружающей среде.
В любом растительном ландшафте, будь то песчаная пустыня Каракумы, или же постоянно влажный экваториальный дождевой лес - гилея, можно наблюдать широкий спектр условий увлажнения - от постоянно влажного до периодически совершенно сухого. Например, в центральной части пустыни Каракумы, сухой бездождевой период, сопровождаемый очень высокими дневными температурами, длится 5-6 месяцев казалось бы, летом высыхает все и до начала осенних дождей какая-либо влага в этом ландшафте отсутствует. Однако на самом деле в продолжение всего знойного лета сухим остается лишь верхний слой песчаного субстрата. На вершинах песчаных гряд глубина просыхания больше, в межгрядовых понижениях - меньше. На определенной глубине влага сохраняется в течение всего года и растения, достигающие своими корнями этого горизонта, в течение всего года обеспечены влагой. В межгрядовых понижениях этот горизонт может находиться всего в 2-3 метрах от поверхности. Благодаря ему в Каракумах когда-то были шире распространены, а в недоступных местах сохранились и доныне, очень своеобразные леса с доминированием черного саксаула.
В экваториальной гилее почва и атмосфера настолько перенасыщены влагой, что перерыв в выпаде! осадков продолжительностью 2-3 недели не оказывает заметного влияния на влагонасыщенность ландшафта. Казалось бы, что ни о какой сухости здесь не может быть и речи, и здесь нет места суккулентным растениям. Однако растения эпифитных местообитаний, которые не имеют контакта с почвой, оказываются в условиях полного отсутствия влаги в окружающей их среде сразу же после того, как просохнет кора дерева-хозяина или же незначительное скопление листового опада в развилке сучьев, то есть уже через несколько часов по выпадения дождя, если речь идет о верхней части кроны. Следовательно, растения эпифиты должны обладать (и обладают) соответствующими механизмами запасания и экономного расходования влаги, чтобы не прекращать жизнедеятельности в перерывах между дождями. Эпифиты отнюдь не ограничиваются в своем распространении дождевыми лесами. Значительно шире последних в тропиках распространены переменно-влажные леса, в которых сухой период может продолжаться 2-3 и более месяцев. Понятно, что распространенные в таких лесах эпифиты являются в высшей мере засухоустойчивыми растениями.
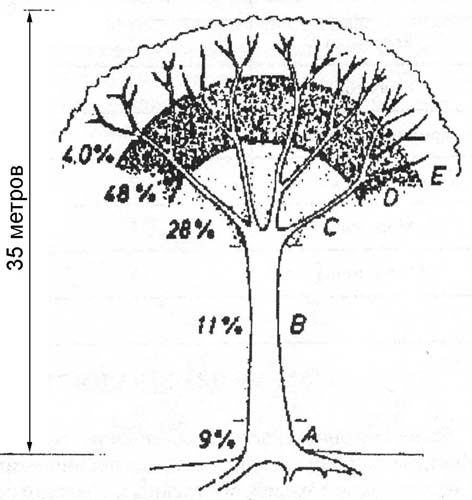
Рис.1. Распространение эпифитных орхидей в кроне дерева-хозяина (по Johanson, 1975) в постоянно влажных дождевых лесах Западной Африки.
Следовательно, мы не вправе ограничивать поиски суккулентных растений лишь "суровыми безводными пустынями". Сухие местообитания распространены повсеместно вместе с древесно-кустарниковой растительностью. Интересно в связи с этим отметить, что значительное количество кактусов также являются эпифитами, например, виды родов Acanthorhipsalis. Aporocactus, Chiapasia, Rhipsalis, Lepismium, Pseudorhipsalts, Hatiora, Erythrorhipsalis, Rhipsalidopsis, Epiphyllanthus, Zygocactus, Pseudozygocactus, Schlumbergera, Marniera и др. Еще более многочисленны эпифитные суккулентные бромелиевые. Но абсолютное первенство среди всех суккулентов вообще по количеству видов принадлежит, вне всякого сомнения, эпифитным орхидеям. Однако с орхидеями связан и самый большой парадокс суккулентологии: суккулентологи не замечают суккулентных орхидей (3).
Цель настоящей статьи - нарушить эту традицию, и дать самые общие сведения по биологии этих интересных растений.
СЕМЕЙСТВО ОРХИДНЫХ (ORCHIDАСЕАЕ A.L.DE JUSSIEU 1789)
Kрупнейшее в классе Однодольных. По разным оценкам, оно включает 600-800 родов и 20-35 тысяч видов. Распространение космополитное, главным образом в тропиках. При этом среди тропических видов доминируют эпифиты (Табл. 1), в время как среди видов умеренных широт их нет совсем и буквально единичные виды эпифитных орхидей заходят в субтропические районы (Флорида, Япония). По данным Madison M. 2) эпифитными являются не менее 20 000 видов орхидей из примерно: 500 родов.
Таблица 1
Распространение орхидей и количество эпифитных видов (по 1, 2, 4, 5, 7)
| Страна, регион
| Количество
родов
| Количество видов
| Из них эпифитных видов
(в %)
|
| Умеренные широты сев. полушария
| 75
| 900
| -
|
| Умеренные широты юж. полушария
| 40
| 500
| -
|
| Канада
| 20
| 68
| -
|
| Германия
| 27
| 53
| -
|
| Территория бывшего СССР
| 30
| 49
| -
|
| Тропическая Америка
| 48
| 152
| -
|
| в том числе: Гватемала
| 306
| 8200
| ?
|
| Венесуэла
| ?
| ?
| 85
|
| Колумбия
| ?
| 1300
| ?
|
| Тропическая Азия
| 250
| 7000
| 7
|
| в том числе: Индия
| ?
| 878
| 61
|
| Малайзия
| ?
| 819
| 73
|
| Филиппины
| ?
| 675
| 81
|
ОСНОВНЫЕ БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ОРХИДЕЙ
Основные биологические особенности орхидей: микотрофность, суккулентность, теневыносливость, пылевидные размеры семян - являются первичными и возникли, как нам представляется, у наземных лилейных предков орхидей, обитавших по берегам пересыхающих водотоков.
Микотрофность
Практически все орхидеи являются облигатными микотрофами и не способны развиваться без контакта с микоризными грибами. У них не развиваются корневые волоски, они продуцируют семена без запаса питательных веществ, неспособные прорасти до тех пор, пока в них не проникнет гифа нужного гриба, от которого орхидея и будет получать все необходимые для роста и развития питательные вещества.
Обычными симбионтами орхидей являются базидиомицеты, главным образом из родов Rhizoctonia, Marasmium, Xerotus, Armillaria, Corticum, Fomes (8). Симбиоз с грибами, которые, в отличие от растений, способны разлагать целлюлозу на моносахариды, дает возможность орхидеям получать полноценное минеральное питание везде, где есть мертвые растительные остатки (опад, кора, мертвая древесина), не дожидаясь их минерализации другими сапрофитными организмами, которыми изобилуют тропические леса: в первую очередь другими грибами, бактериями, актиномицетами и членистоногими (термиты, свободно живущие нематоды, клещи, ногохвостки, личинки мелких двукрылых, личинки и взрослые особи мелких жуков, трипсы, тли, сеноеды, детритоядные многоножки и др.). Таким образом, орхидеи, вследствие микотрофносги, являются консументами дерева-хозяина, получая полноценное минеральное питание из мертвых продуктов его жизнедеятельности.
Микроскопический размер семян

Epipogium aphyllum
Семена орхидей в целом очень мелкие, меньше семян других цветковых растений, что характерно для большинства видов орхидей, как эпифитных, так и наземных. Зародыш в семени не дифференцирован и сильно редуцирован, во многих случаях до нескольких десятков клеток (у Epipogium aphyllum зародыш в зрелом семени состоит всего из 9 клеток). Запас питательных веществ отсутствует. Каждый плод содержит многие тысячи, и даже сотни тысяч семян, у некоторых видов Cattleya один плод может содержать 4 миллиона семян (Поддубная-Арнольди, 1954). Микроскопические размеры семян позволяют эпифитным орхидеям беспрепятственно расселяться в нужном ярусе леса.
Эволюционная тенденция в сторону уменьшения массы одного семени и увеличения количества семян в плоде возникла, как нам представляется, еще у наземных предков орхидей как реакция на низкую биологическую продуктивность вследствие экстремальных условий существования. Возникшие симбиотические взаимоотношения с грибами усилили эволюцию в этом направлении, т.к. исчезла необходимость запасать в семенах питательные вещества. Дополнительным стимулом послужила адаптация орхидей к эпифитному образу жизни.

Cattleya sp. (link)
Суккулентность
Выше отмечалось, что кора ветвей и стволов деревьев-хозяев, как и незначительные скопления опада в развилках ветвей, являющегося субстратом для питающих корней, не могут, в отличие от почвы, накопить сколько-нибудь значительного запаса влаги. Десятидневный перерыв в выпадении атмосферных осадков мало влияет на влагосодержание лесных почв. В то же время двухдневный перерыв полностью высушивает эпифитные местообитания тропического леса. Поэтому эпифитные орхидеи (и не только они) вынуждены иметь эффективные механизмы регулирования водного баланса.
Эффективность механизмов регулирования водного баланса должна возрастать как с увеличением продолжительности сухого сезона, так и с занятием более высокого яруса в кроне дерева. Исследования микроклимата различных вариантов тропического леса показали, что минимальные колебания температуры и влажности воздуха, известные нам из популярных описаний, присущи только приземному ярусу.
С повышением яруса резко возрастает амплитуда суточных колебаний основных метеорологических параметров, таких, как температура и влажность воздуха, освещенность, скорость воздушных потоков и др. (Г. Вальтер "Рас-тительность Земного шара", т. 1, 1968). В верхней наружной части древесной кроны даже в постоянно влажных дождевых экваториальных лесах - гилеях относительная влажность воздуха в отдельные дневные часы может понижаться до 50%, что в принципе невозможно в приземном ярусе. Повышенные скорости ветра в верхнем ярусе кроны еще больше способствуют иссушению этого биотопа. И, хотя понижения относительной влажности воздуха до указанных выше значений во влажнотропическом лесу кратковременны и продолжаются не более нескольких часов, тем не менее, даже древесные растения испытывают в это время жесткий дефицит влаги. И совсем не случайно у деревьев верхнего яруса листья очень часто имеют ксероморфную структуру.
Большинство эпифитных видов орхидей суккулентны: их фотосинтезирующие органы содержат специализированные водозапасающие клетки, а фотосинтез осуществляется по САМ-типу. Насколько необходим САМ-тип фотосинтеза эпифитным орхидеям, показывают результаты следующего опыта по высушиванию орхидей, проводившегося в Сингапуре (C.J.Goh, 1981): три вида орхидей: Eria velutina, Dendrobium tortile и D. cruminatum были помещены под навес, исключавший попадание на растения влаги атмосферных осадков. Все три вида содержали примерно равное количество воды - 96-98%, однако Eria velutina транспирировала по обычному типу, а дендробиумы - по суккулентному. За 20 дней содержание воды в Eria velutina снизилось с 98% до 65%. В суккулентных дендробиумах содержание воды через 20 дней осталось выше 90%.

Eria velutina (link)
Так же, как среди других суккулентных растений, среди орхидей есть и листовые, и стеблевые, и смешанные формы суккулентности. У орхидей с симподиальной формой роста (а их подавляющее большинство) основными водозапасающими органами являются псевдобульбы - утолщенные побеги, образовавшиеся из одного или нескольких междоузлий и имеющие самые разнообразные объемные формы: луковицеобразные, шаровидные, веретенообразные и др. Размер их различен: от нескольких миллиметров в диаметре у карликовых безлистных бульбофиллюмов до 15 см в диаметре у Peristeria elata. Псевдобульба может нести один или несколько листьев на верхушке, или на поверхности псевдобульбы. Внутренняя часть псевдобульб заполнена водозапасающими клетками.

Dendrobium tortile (фото Eric Hunt, link)
Если в течение продолжительного сухого периода орхидея сбрасывает листья (одним из механизмов регулирования водного баланса орхидей служит слой клеток в основании листьев, предназначенный для сбрасывания листьев в сухой сезон), фотосинтез продолжает осуществляться только поверхностью псевдобульб. Однако газообмен с атмосферой осуществляется не клетками эпидермиса, как у "обычных" суккулентов, а через углубление в верхней части псевдобульбы, поверхность которого усеяна многочисленными устьицами. Это принципиально отличает орхидеи от других стеблевых суккулентов. Объяснить этот феномен можно тем, что орхидеи почти никогда не подвергаются перегреву солнечными лучами и, таким образом, у них отсутствует необходимость борьбы с перегревом фотосинтезирующих органов путем интенсивной транспирации.
У орхидей с моноподиальной формой роста, например, у видов Vanilla, у которых стебель не образует псевдобульб, фотосинтезирующие клетки функционируют по обычному для суккулентов типу.
Листья орхидей, простые и цельные с параллельным жилкованием, почти всегда утолщенные за счет клеток водозапасающей паренхимы, также очень разнообразны и могут являться основными водозапасающими органами.
Эпифитные орхидеи могут также запасать воду в корнях, однако это принципиально иной тип водозапасания, нежели суккулентность: наряду с питающими корнями, "зараженными" грибом, которые внедряются в субстрат и обеспечивают минеральное питание растений, эпифитные орхидеи развивают также воздушные висячие корни, покрытые слоями мертвых пустых клеток - веламеном. В перенасыщенной влагой атмосфере слои веламена способны поглощать водяной пар и влагу атмосферных осадков, впитывая их подобно губке и сохраняя какое-то время. В менее влажных условиях значение веламена снижается и возрастает значение суккулентности.
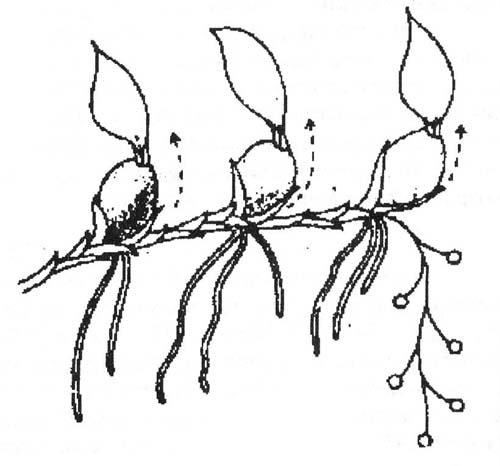
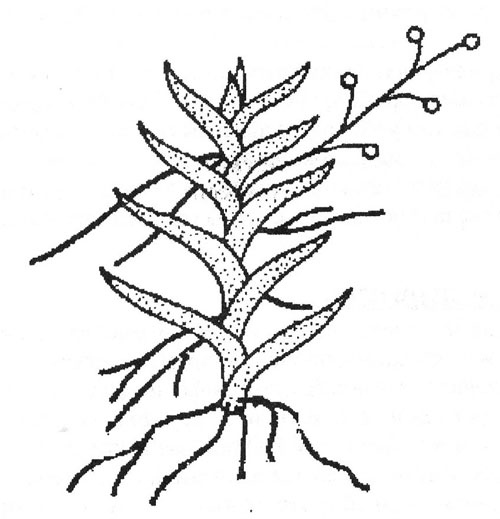
Рис. Симподиальная и моноподиальная форма роста орхидей (по Kluge)
Теневыносливость
Большинство орхидей характеризуются высокой теневыносливостью, во многих случаях максимальной среди цветковых растений. По данным исследований, проводившихся в дождевых лесах Западной Африки (Johanson, 1975.цит.по 1) 9% видов эпифитных орхидей, из учтенных 101 вида, занимали базальные части стволов деревьев до высоты 3 м над уровнем почвы. В этом ярусе условия освещения экстремальные для цветковых растений, т.к. освещенность ниже 1% от полной. 11% видов занимали средние части стволов от 3 м и до развилки основных сучьев. 28% были расселены на нижней трети основных сучьев, 48% - на средней трети основных сучьев, и 4% - на внешней трети основных сучьев. Все виды орхидей в этом лесу притенялись листвой концевых ветвей, но орхидеи внешней трети основных сучьев (4%) в течение дня попадали под прямые лучи экваториального солнца.
Несомненно, что высокая теневыносливость орхидей обусловлена их микотрофностыо. Однако это не дает повода говорить о принципиальном отличии орхидей от других суккулентов, т.к. очень многие "типичные" суккуленты также избегают открытых пространств и поселяются в подкроновом пространстве ксерофильных деревьев и кустарников. Это особенно характерно для представителей подсемейства Stapelieae (White A. & Sloane B.L.: The Stapelieae. Ed.I., Pasadena 1934). Исключительная тенелюбивость указывается для кактуса Obregonia denegrii Fric (J.Riga, R.Subik: Nekolik poznamek k ekologii Obregonia denegrii Fric-Kaktusy (2) 34-36, 1978.).
Только в тени растут эпифитные виды кактусов.

Vanilla planifolia (link)
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ОРХИДЕЙ
Особенности биологии орхидей дают основание предполагать, что предки этих растений первоначалъно заселяли переменно-влажные местообитания, характеризовавшиеся большим количеством мертвых растительных остатков и высокой затененностью.
Облигатная микотрофность орхидей свидетельствует об очень долгом эволюционном развитии в таких условиях, так как факультативная микотрофность в растительном мире является едва ли не универсальным правилом. Кроме того, в постоянно влажных условиях, например, в гилеях, отсутствуют условия для накопления мертвых растительных остатков. Для немикотрофного растения эпифитные местообитания непригодны для жизни в силу бесплодности субстрата. Наоборот, для микотрофного растения кора и древесный опад растения-хозяина являются полноценным субстратом.
Можно предположить, что когда предки орхидей проникли в постоянно-влажные экваториальные леса, они не смогли заселить наземный ярус вследствие как отсутствия подстилки, так и чрезмерной даже для орхидей затененности, но легко проникли в верхний ярус, обладая суккулентной физиологией и пылевидными семенами. Здесь, в благоприятных условиях питания, увлажнения и освещения они дали вспышку видообразования.
Таким образом, мы можем сделать вывод, что эпифитные суккулентные орхидеи по основополагающим морфологическим, физиологическим и экологическим признакам аналогичны наземным суккулентам. Мезофилл фотосинтезирующих органов разделен на хлоренхиму и водозапасающие паренхимные клетки, фотосинтез осуществляется по САМ-типу, растения занимают биотопы с периодически полным отсутствием влаги в окружающей среде. Именно суккулентность эпифитных орхидей позволяет понять причины их полного отсутствия в лесах умеренного и почти полного - в лесах субтропического климатических поясов: водозапасающие органы неустойчивы по отношению к отрицательным температурам, а для несуккулентных орхидей эти биотопы экстремально сухие.
В то же время эпифитные орхидеи имеют много своеобразных черт: облигатная микотрофность, теневыносливость. Уникальной для суккулентов является морфология симподиальных орхидей, а также особенности их газообмена.
ЛИТЕРАТУРА
- CJ.Goh & Kluge M. - Gas exchange and water relations in Epiphytic Orchids in Vascular Plants Epiphyte. Ulrich Luttge (ed.) Berlin, 1989, pg. 139-166.
- Madison M. Vascular Epiphytes: their systematic occurrence and salient features. Selbyana 2:1 -13,1977
- IOS, - Repertorium Plantamm Succulentarum. Zurich. XLI-XLIV.
- Dressier R.L,- The orchids. Natural history and classification. Harvard University Press. Cambridge. Massachusetts, 1981.
- Lang D. - Orchids of Britain. Oxford University Press. Oxford, 1980.
- Вальтер Г. - Растительность Земного шара. "Прогресс", М., 1968.
- Вахрамеева М.Г. идр. - Орхидеи нашей страны. М."Наука", 1991
- Терехин Э.С. - Паразитные цветковые растения. Л.,"Наука", 1977






