ЖУРНАЛ "КАКТУСЫ И ДРУГИЕ СУХОЛЮБИВЫЕ РАСТЕНИЯ" - выпуск 15-16 (1-2 за 2003 год)
РОДОВЫЕ ОТНОШЕНИЯ В Alooideae (Asphodelaceae)
Гидеон Ф. Смит и Бен-Эрик Ван Вик (Журнал “Taxon”, вып.40, ноябрь 1991)
Smith, Gideon F. & van Wyk, Ben-Erik: Generic relationships in the Alooideae (Asphodelaceae). – Taxon 40: 557-581. 1991.
ОТ РЕДАКЦИИ. Статья переведена и подготовлена к печати Андреем Константиновичем Шереметом – руководителем секции любителей Алоэ РОИКС. Она публикуется с любезного разрешения авторов. Редколлегия благодарит Гидеона Ф. Смита и Бен-Эрик Ван Вика за предоставленные материалы и любезное разрешение опубликовать их на страницах нашего журнала. Editorial Staff thanks Smith, Gideon F. & van Wyk, Ben-Erik very much for the given materials and kind Sanction to publish it.
АБСТРАКТ. В семействе Asphodelaceae, подсемейство Alooideae - естественная (монофилетическая) группа. По современным воззрениям систематиков, подсемейство Alooideae выделяется в самостоятельное семейство Aloaceae (G.F.Smith & L.E.Newton, ALOACEAE, цит.по: Eggli, Urs (ed.) 2001. Monocotyledons – В: U.Eggli (ed.), Illustrated handbook of succulent plants). Следовательно, на основе обладания характерными признаками, отличающими эту группу от остальных родов Asphodelaceae, все рода этого подсемейства могут рассматриваться, как происходящие от общего предка. Невозможно, однако, всесторонне интерпретировать филогенетические отношения в подсемействе, основываясь лишь на сравнительно-морфологических данных и фитогеографических соображениях.
В настоящей работе филогенетические отношения родов Aloe, Astroloba, Chortolirion, Gasteria, Haworthia, Lomatophyllum и Poellnitzia исследуются с использованием кладистической методологии. Наиболее очевидные общие признаки этих родов – суккулентная консистенция листьев, серповидный или ладьевидный в поперечном разрезе контур листа и выраженный бимодальный кариотип 2n = 14 хромосом. Для кладистического анализа использовались 16 отличительных признаков; род Kniphofia был выбран как вне групповой на основе двух синапоморф с Alooideae, а именно – трубчатые цветки и сросшиеся доли околоцветника. Признаки подробно обсуждены, поляризованы по принципу плезиоморфности – апоморфности состояния, обработаны при помощи пакета программ "Hennig86" и затем использованы для построения филогенетической гипотезы.
Введение
Подсемейство Alooideae (Asphodelaceae) по Dahlgren & al. (1985) включает 7 родов и приблизительно 450 видов (Табл. 1). Преимущественно это группа Старого Света, большинство же родов встречается в Африке южнее Сахары. Род Aloe найден также на Аравийском полуострове, Мадагаскаре и Сокотре, в то время, как Lomatophyllum был описан на островах Альдабра, Мадагаскар и Маврикий. Самая большая концентрация родов и видов - в южной Африке, то есть, грубо говоря, в регионе к югу от рек Кунене, Окаванго и Лимпопо. В этом регионе Liliaceae, в которое были изначально включены алоидные таксоны, представляет основную часть лепестковых однодольных и составляет существенную часть флоры (Goldblatt, 1978; Gibbs Russell, 1985, 1987).
Таблица 1. Роды Asphodelaceae по Dahlgren & al. (1985)
| Таксоны | Число видов | Географическое распространение | Источники данных
|
| Asphodeloideae | | |
|
| Asphodeline Reichenb. | 14 | Средиземноморье, Западная Азия | Tuzlaci (1987)
|
| Asphodelus L. | 12 | Средиземноморье, Западная Азия | Dahlgren & al. (1985)
|
| Bulbine Wolf | 60 | Южная Африка, Австралия | Watson (1987)
|
| Bulbinella Kunth | 16 | Южная Африка, Новая Зеландия | Perry (1987)
|
| Bulbinopsis Borzi | | - однородный с Bulbine | Watson (1986a)
|
| Eremurus M. Bieb | 35 | Центральная Азия | Dahlgren & al. (1985)
|
| Jodrellia Baijnath | 3 | Тропическая Африка | Baijnath (1978)
|
| Kniphofia Moench | 70 | Ю.и В.Афр., Мадагаскар, Ю.Арав. | Codd (1968) Marais (1973)
|
| Simethis Kunth | 1 | Зап. Европа, Зап. Средиземноморье | Dahlgren & al. (1985)
|
| Trachyandra Kunth | 50 | Южная Африка | Obermeyer (1962)
|
| Verinea Pomel | | - однородный с Asphodelus | Jackson (1894-1895)
|
| Alooideae | | |
|
| Aloe L | 333 | Южн. и В.Афр., Мад-р, Южн.Арав. | G.Reynolds (1966, 1982)
|
| Astroloba Uitew. | 7 | Юж. Африка (финбош, сукк. кару) | R. Reinecke (1965)
|
| Chortolirion A. Berger | 1 | Южн. Африка (гл. обр. прерия) | Smith (1988)
|
| Gasteria Duval | 14 | Южн. Африка (гл. обр. финбош) | Van Jaarsveld (1989)
|
| Haworthia Duval | 68 | Южн. Африка (гл. обр. финбош) | Bayer (1982)
|
| Lomatophyllum Willd. | 12 | Мадагаскар, Маврикий | Jacobsen (1986)
|
| Poellnitzia Uitew | 1 | Ю. Африка (сукк. кару) | Smith (1985)
|
Большинство исследователей считает Alooideae таксономически трудным подсемейством. Морфология листа суккулентов часто значительно изменяется в зависимости от условий окружающей среды (ср. Schelpe, 1958, и Bayer, 1980 на Gasteria Duval) и, в некоторых случаях, таксоны кажутся гибридами (ср. Bayer, 1975 и Glen & Hardy, 1987 на Aloe серии Saponaria А. Berger; Bayer, 1973 и Scott, 1981 на Haworthia reinwardtii Haw. - H. coarctata Haw.). Недавние исследования, преимущественно таксономического характера, особенно на родах Aloe, Gasteria и Haworthia, повлекли изучение особенностей морфологии (G. W. Reynolds, 1966, 1982; Van Jaarsveld, 1989; Bayer, 1982; Scott, 1985), химии экссудата листа (обозрение T. Reynolds, 1985a, b; ср. T. Reynolds, 1990), рельефа поверхности листа (ср. Cutler, 1972, 1982) и цитогенетики (ср. Riley и Majumdar, 1979). Более того, за исключением последних работ Smith (1988, 1990, 1991), мелкие роды Alooideae (Таблица 1) были опущены из большинства предыдущих таксономических и биосистематических исследований этой группы (см. также Beaumont и др., 1985). Начиная с первых, работы на крупных родах были связаны с основной проблемой паттернов изменчивости и лучшему их таксономическому опознанию, и лишь незначительное внимание было уделено возможным филетическим отношениям. Фенетически роды Alooideae недавно анализировались в численно-таксономических работах Rowley (1967a, 1969). Сомнительный таксономический статус некоторых из признанных родов делает результаты этих исследований неубедительными (Bayer, 1972; Rowley, 1976a; Ivimey-Cook, 1971). Что касается нескольких предыдущих работ по эволюции отдельных родов Alooideae (Kok, 1973; Holland, 1978), то они часто высоко спекулятивны и явно конъюнктурны (Newton, 1973). Представленный кладистический анализ позволяет лучше понять взаимоотношения членов Alooideae и впервые сформулировать гипотезу филогенетических отношений в пределах подсемейства.
МАТЕРИАЛЫ И МЕТОДЫ
Роды Alooideae, признанные в этой работе. – Подсемейство Alooideae, Asphodelaceae, традиционно трактовалось как одна из 28 триб Liliaceae, а именно Aloeae (Hutchinson, 1959). Позднее были опубликованы две обширные ревизии систем классификации (Cronquist, 1981; Dahlgren & al., 1985), которые не только отличаются от системы Hutchinson (1959), но и друг от друга. Cronquist (1981) включил алоидные рода, наряду с Kniphofia Moench, в Aloaceae. Dahlgren & al. (1985) выделили алоидные рода в подсемейство Alooideae, Asphodelaceae, тогда как род Kniphofia был отнесен ими к Asphodeloideae. Приблизительно 450 видов, включенных в Alooideae, объединялись в 27 родов. В настоящее время общепризнанны только семь из этих родов. Для целей настоящей статьи принята следующая родовая синонимия (о дополнительной номенклатурной информации относительно синонимов следует справляться у Rowley, 1976 a, b. Для каждого рода даются ссылки, недавно дополненные):
- Aloe Linnaeus (1753: 319; ср. G. W. Reynolds, 1966, 1982). Син.: Catevala Medikus (1786: 67) частично; Kumara Medikus (1786: 69); Rhipidodendrum Willdenow (1811: 164); Pachidendron Haworth (1821: 35); Bowiea Haworth (1824: 299) и не J. D. Hooker (1867: t. 5619); Agriodendron Endlicher (1836-1840: 144); Papilista Rafinesque (1840: 137); Succosaria Rafinesque (1840: 137); Busipho Salisbury (1866: 76); Ptyas Salisbury (1866: 76); Chamaealoe A. Berger (1905: 43); Leptaloe Stapf (1933: t. 9300); Aloinella Lemee (1939: 27) не Cardot (1909: 76); Guillauminia Bertrand (1956: 41).
- Gasteria Duval (1809: 6; ср. Van Jaarsveld, 1989). Син.: Atevala Rafinesque (1840: 136).
- Haworthia Duval (1809: 7) сохр. назв. (ср. Bayer, 1982). Син.: Catevala Medikus (1786: 67) частично; Apicra Willdenow (1811: 167), а не Haworth (1819: 61); Kumaria Rafinesque (1840: 137); Tulista Rafinesque (1840: 137).
- Lomatophyllum Willdenow (1811: 166; ср. Jacobsen, 1986). Син.: Phylloma Ker (1813: t. 1585).
- Chortolirion Berger (1908: 72; ср. Smith, 1985).
- Poellnitzia Uitewaal (1940: 61; ср. Smith, 1985).
- Astroloba Uitewaal (1947a: 53; ср. Roberts Reinecke, 1965). Син.: Apicra Haworth (1819: 61) не Willdenow: 167 (1811).
Эти семь родов были приняты за единицы изучения (ЭЕ, Эволюционные Единицы, fide Estabrook, 1977), но предположение о монофилитичности родов возникает как основная проблема традиционного определения границ родов, особенно по отношению к Aloe и Haworthia.
Используемые данные. - Источники данных, используемых в кладистическом анализе, приведены в Таблице 1. Дополнительная информация была получена из Coetzee и Van der Schijff (1969), Baijnath (1980) и Van der Riet (1977). Сахароидный состав нектара был проанализирован одним из нас (B-E. Van Wyk). Хромосомные данные получены, главным образом, от De Wet (1960), Sharma и Mallick (1966), Jones и Smith (1967), Brandham (1971) и Riley и Majumdar (1979). Источники дистрибутивной информации приведены в надписях на Fig. 3-7.
Все признаки таксонов потенциально полезны для выявления эволюционных тенденций. Однако лишь некоторые из них являются достаточно надежными индикаторами эволюционной направленности для использования в кладистическом анализе. В настоящей работе использованы 16 признаков (Табл. 2), т.е. все те, примитивность или продвинутость которых мы могли с уверенностью определить. Анатомические признаки были исключены, несмотря на их потенциальную ценность для подтверждения монофилитичности Alooideae и различия между последним и Kniphofia. Рельеф поверхности листа и типы пучковых влагалищ особенно важны в этом отношении (Д-р D. F. Cutler, личное сообщение), однако включение этих признаков должно основываться на более тщательном сравнении всех родов. Матрица основных данных показана в Табл. 3.
Таблица 2. Постулированный ряд состояний 16 признаков, используемых для построения кладограммы на рис. 2.
| | Отличительный признак | Установленные состояния
|
| 1 | Характер произрастания | бесстебельное – 0; стебельное – 1
|
| 2 | Расположение листьев | густое – 0; рыхлое – 1
|
| 3 | Консистенция листа | мезофитная – 0; суккулентная – 1
|
| 4 | Бугорчатость листа | отсутствует – 0; имеется – 1
|
| 5 | Профиль поперечного сечения листа | V-видный – 0; серповидный/ладьевидный – 1
|
| 6 | Плотность соцветия | густое – 0; рыхлое – 1
|
| 7 | Форма цветка | не трубчатая – 0; трубчатая – 1
|
| 8 | Слияние долей околоцветника | свободные – 0; сросшиеся – 1
|
| 9 | Окраска цветка | жёлтая – красная – 0; зеленая - белая – 1
|
| 10 | Положение цветка | ненаклонное вверх – 0; наклонное вверх – 1
|
| 11 | Симметрия околоцветника | не двугубый – 0; двугубый – 1
|
| 12 | Цветочный аромат | имеется – 0; отсутствует – 1
|
| 13 | Положение пыльника | снаружи – 0; внутри – 1
|
| 14 | Сахара нектара | сахароза почти отсутствует – 0; сахароза преобладает – 1
|
| 15 | Сахара нектара | глюкозы и фруктозы поровну – 0; глюкозы много больше –1
|
| 16 | Основное число хромосом (x) | 6 – 0; 7 – 1
|
Таблица 3. Филогенетические отличительные признаки родов Kniphofia (Kni), Aloe (Alo), Astroloba (Ast), Chortolirion (Cho), Gasteria (Gas), Haworthia (Hw), Lomatophyllum (Lm) и Poellnitzia (Poe).
Нумерация признаков аналогична табл. 2. Hyp – гипотетический предок. Отличительные признаки: 0 – плезиоморфный; 1 – апоморфный; ? – сомнительный признак, данные не доступны.
| номера признаков | | | | | | | | | | | | | | | |
|
| tax | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16
|
| Hyp | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0
|
| Kni | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0
|
| Alo | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1
|
| Ast | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1
|
| Cho | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1
|
| Gas | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1
|
| Hw | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1
|
| Lm | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | ? | 0 | 0 | 0 | 1
|
| Poe | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1
|
Поляризация состояний признаков. — Выбор оперативных критериев для определения примитивных (плезиоморфных) и продвинутых (апоморфных) состояний признаков – один из наиболее трудных, но важных шагов к познанию филогении (ср. Cronquist, 1987). В виду отсутствия окаменелостей в подсемействе Alooideae и относительно единообразного бимодального кариотипа его членов (x=7; одна длинная субметацентрическая, три длинных и три коротких акроцентрических; ср. Brandham & Johnson, 1977; и Brandham, 1983), разветвленная модель эволюции этой группы была построена путем оценки эволюционного значения состояний признаков существующих таксонов. Поляризация состояний признаков была выполнена по отношению к выбраному внегрупповому таксону (Maddison & al., 1984) – в данном случае роду Kniphofia. Moнофилия Kniphofia и родов из Alooideae демонстрируется двумя синапоморфами, а именно: трубчатый околоцветник (признак 7 в табл. 2) и срастание околоцветника (признак 8 в табл. 2).
Используемый кладистический метод. - Основная методология, которой мы следовали, это кладистический или филогенетический анализ, описанный Humphries и Funk (1984) и Linder (1988). Вышеупомянутая матрица данных (Таблица 3) была проанализирована с использованием пакета программ "Hennig86", который является методом, выводящим филогению по принципу максимальной экономии.
Результаты и обсуждение
Эти 16 признаков и их состояния (Таблица 2), используемые в матрице основных данных (Таблица 3), внесены в список и обсуждены ниже. В настоящем анализе межродовых отношений в пределах Alooideae сделана попытка скомбинировать всю доступную информацию, поддающуюся филогенетической интерпретации. Эти данные приведены в виде кладограммы (рис. 2). При использовании "ie" опции "Hennig86" данные ведут к единственной истинной кладограмме с последовательностью коэффициентной величины 88 и протяженностью длиной 18 состояний признаков изменений.
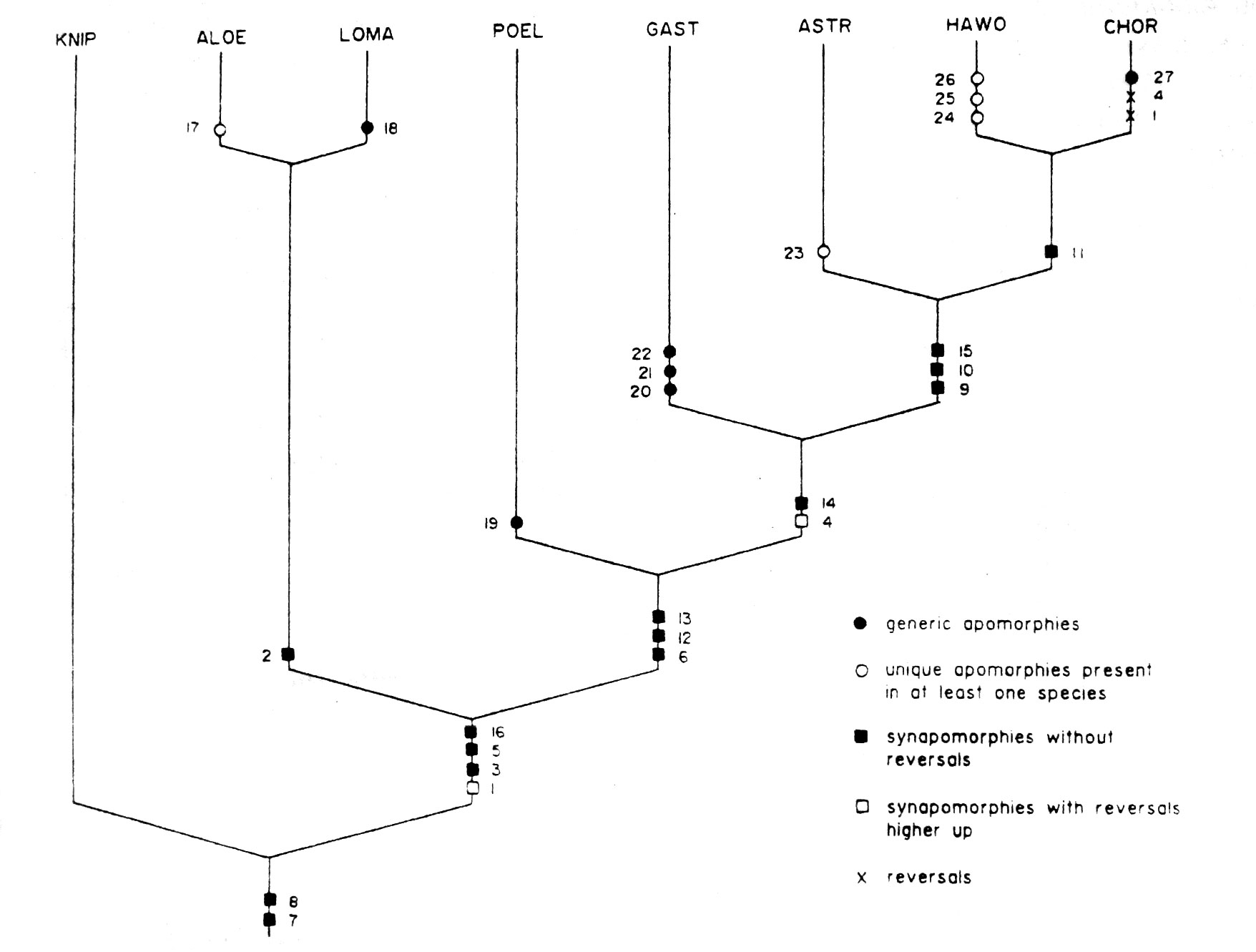
Рис 2. Кладограма предполагаемых филогенетических связей между родами Aloe, Astroloba, Chortolirion, Gasteria, Haworthia, Kniphofia, Lomatophyllum и Poellnitzia. Значения №№ 1-16 приведены в Табл. 2. Наличие тех или иных признаков приведено в Табл. 3.
|
1. Характер произрастания
Подсемейство Alooideae включает таксоны, демонстрирующие широкий диапазон жизненных форм, начиная с геофитов (Chortolirion), небольших высокоспециализированных розеточных суккулентов (например, Aloe [секция Aloe, подсекция Humiles Berger], Astroloba, Haworthia), кустарников (например, мадагаскарские кустарниковые виды алоэ, типа A. acutissima Perr.) и вьющихся растений (Aloe ser. Macrifoliae Haw. [секция Aloe, подсекция Prolongatae Berger]), и до небольших или крупных деревьев (например, Aloe секции Dracoaloe Berger). На основе определительных ключей Aloe G. W. Reynolds-а (1950a, 1966), Holland (1978) допустил, что древовидные формы этого рода (к которым относится примерно13% видов) более продвинуты, нежели травянистые формы. Примечательно, что Reynolds не поддерживал эту точку зрения, поскольку он интерпретировал A. suzannae Decary (самый высокий из малагасийских алоэ с высотой стебля до 3-4 м) как наиболее примитивный вид (G. W. Reynolds, 1966). Предположение Holland-а(1978) не поддержано и цитогенетическими данными, представленными Brandham (1983), предположившим, что виды алоэ с лазающим характером произрастания и обычно сравнительно мезофитными листьями, типа A. tenuior Haw. и A. ciliaris Haw. (Hunt, 1978), представляют примитивное состояние. Мы в значительной степени поддерживаем точку зрения Brandham (1983) и предполагаем, что Alooideae (включая как мелкие высоко суккулентные таксоны, так и древовидные формы) происходит от мезофитного, относительно бесстебельного таксона.
Kamstra (неопубл.) также доказывает, что предок алоэ был маленьким растением, вероятно около 150 мм высотой. Это предположение, во-первых, основано на том факте, что и в других родах подсемейства Alooideae имеются виды относительно небольшой высоты, например, Astroloba, Chortolirion и Haworthia. Во-вторых, это справедливо и для членов близкородственного подсемейства Asphodeloideae (триба Asphodeleae по Hutchinson, 1959), которые достигли относительно широкого расселения в Гондване, например, Bulbine Wolf, которая встречается и в Австралии (Watson, 1987), и Bulbinella Kunth, имеющая афро-новозеландский ареал (Watson, 1986a, b, 1987; Perry, 1987, 1990).
В роду Kniphofia есть один или два вида (типа K. northiae Baker), изредка способных развить надземный стебель более 0.5 м. длиной (Roux, 1985). Однако, подавляющее большинство видов Kniphofia - бесстебельные (Codd, 1968). По отношению к внегрупповой Kniphofia, Chortolirion, имеющий короткую вертикальную ось со слабомясистыми основаниями листьев, выглядит примитивным (a reversal). Среди изменчивых Aloe также имеется несколько бульбоидных видов, например, A. buettnerri A. Berger (Jankowitz, 1975) и A. kniphofioides Baker (Dyer, 1951). Однако, у более чем 90 % видов алоэ развивается стебель, хотя порой и очень короткий (например, A. juvenna Brandham и Carter, 1979). Стебельные виды встречаются и во всех других таксонах Alooideae, особенно Astroloba, Haworthia subg. Hexangulares Uitew. ex М. Bayer, Lomatophyllum и Poellnitzia. В роде Gasteria немногие стебельные виды (G. rawlinsonii Oberm. и G. bicolor Haw.) являются скорее исключениями из правила. У них формируются длинные облиствленные стебли вместо прикорневых (концентрических или двурядных) розеток (Obermeyer, 1976). Примечательно, что древовидный или кустарниковидный характер произрастания независимо развился и в других таксонах однодольных, таких как Yucca L., Agavaceae (Dahlgren и Clifford, 1982).
Вопреки ожиданиям, что вторичное утолщение может быть связано с характером произрастания растений, эти признаки не полностью коррелируют в подсемействе Alooideae. Так, Coetzee и Van der Schijff (1969) показали, что геофитные асфоделовые таксоны, типа Chortolirion stenophyllum (Baker) А. Berger, Bulbine coetzeei Oberm. и Trachyandra saltii (Baker) Oberm.,– все имеют вторичное утолщение, которое, однако, у них явно много меньше, чем у растений известных древовидных таксонов, например, Aloe (G. W. Reynolds, 1950b; Cutter, 1971; Esau, 1977; Jankowitz, 1985; Rebelo и др., 1989). Вторичное утолщение также встречается и у Kniphofia (Dahlgren и др., 1985). Как и в случае с развитием древовидности, способность к вторичному утолщению, вероятно, возникала в эволюции неоднократно и развилась независимо в нескольких различных семействах однодольных, таких как Asphodelaceae (Dahlgren и Rasmussen, 1983), Iridaceae (Rudall, 1984, 1989; Manning и др., 1990) и Dracaenaceae (Dahlgren и др., 1985.). Распространение этого признака в Alooideae недостаточно изучено, чтобы использовать его как родовую синапоморфу.
2. Расположение листьев
В основном, двусторонние листья большинства таксонов Alooideae сильно пучкуются на вершине (древовидные виды) или в прикорневых розетках. Не суккулентные листья книфофии обычно густо растут в прикорневых розетках и расположены в отчетливых рядах. Кроме того, листья большинства таксонов Alooideae, за исключением Aloe и Lomatophyllum, тесно сжаты, плотно располагаясь на облиствленном стебле или в прикорневой розетке. В родах же Aloe и Lomatophyllum есть виды, листья которых расставлены сравнительно широко. Несколько оконцелистных видов есть только в роде Haworthia (Schonland, 1910a, b; Hutchison, 1951; Rauh, 1974; Krulik, 1980; Cutler, 1985) и, следовательно, этот признак неинформативен для выявления родовых отношений. Равным образом, неотеническое сохранение двурядности листьев развилось у отдельных видов как Aloe, так и Gasteria (Takhtajan, 1976; Rauh, 1977).
3. Консистенция листьев
Alooideae полностью состоит из листовых суккулентов. Предположительно, листовая суккулентность таксонов Alooideae является характерной специализацией, развившейся в ответ на воздействие аридности в комбинации с огнем (пожаром) (Laubscher, 1973a; Thomas и Goodson, 1986; Scholes, 1988; Weisser и Deall, 1989; но см. Bond, 1983 на Aloe ferox Miller, и Van Jaarsveld, 1987 на A. plicatilis (L.) Miller) и других переменных окружающей среды типа высоких уровней освещенности (Pearson, 1914; Lewis и Nobel, 1977; Eller и др., 1983; Larcher, 1983; Cutler, 1985; Nobel, 1989).
Некоторые таксоны Alooideae, встречающиеся в прериях (часто с высоким уровнем осадков), особенно алоэ секции Graminialoe G. Reyn. и алоэ секции Leptoaloe A. Berger (Laubscher, 1973a, b), и Chortolirion, похоже, вернулись к менее суккулентной консистенции листьев. У большинства этих видов – сокращающиеся корни и наблюдается тенденция к геофитности.
4. Бугорчатость листьев
Предположительно, наличие характерных белых или зеленых сосочков на листьях некоторых видов Astroloba, Gasteria и Haworthia является наследуемым состоянием. Редкая встречаемость этого признака у алоэ (например, A. aristata Haw.) рассматривается здесь, как результат параллелизма. Этот признак, возможно, развился в ответ на прессинг травоядных. Листья Gasteria особенно пользуются спросом у травоядных (Van Jaarsveld, 1989), поскольку в них отсутствует горький ингредиент, найденный в листьях алоэ (Watt и Breyer-Brandwijk, 1962). Наличие осязаемых, твердых сосочков, часто сливающихся в полосы, делает, вероятно, листья менее аппетитными для животных и может представлять форму механической защиты. У видов рода Gasteria листовые сосочки особенно заметны в ювенильном состоянии, листья же взрослых экземпляров часто бывают гладкими, например, у G. acinacifolia (Jacq). Haw. Обычно, в таких случаях листья взрослых экземпляров обильно покрыты пятнами, по-видимому, как камуфляж от травоядных. Во-вторых, подобно рефлексивной функции поверхностного слоя частиц воска или волосков, способных увеличивать отражение от листьев листового суккулента Cotyledon orbiculata L. на 22 % (в видимом спектре) и на 12 % (в инфракрасном спектре) (Sinclair и Thomas, 1970), сосочки могут быть адаптацией, предохраняющей ткани, лежащие в глубине листа, от избыточной освещенности и от термических повреждений. Интересно, что бугорчатость листьев является доминантным признаком, как показали первые же возвратные скрещивания между гладколистной Haworthia cymbiformis (Haw.) Duval и видом с рельефными сосочками на листьях – H. attenuata Haw. Отсутствие листовых сосочков у Chortolirion и некоторых видов Haworthia рассматривается здесь как реверсия.
5. Профиль поперечного сечения
В поперечном сечении несуккулентные листья видов Kniphofia отчетливо килевидные (V-видные), за исключением некоторых клонов K. northiae и K. stricta Codd (Codd, 1968; Baijnath, 1980). Напротив, поперечное сечение листьев большинства видов Alooideae серповидное или ладьевидное. Последний контур рассматривается как результат эволюционирования листьев с суккулентной консистентностью (см. 3, выше). Серповидный или ладьевидный профиль - очевидная адаптация к засушливости и представляет синапоморфу для всех таксонов Alooideae.
Многие виды Alooideae демонстрируют на нижней стороне листьев обычно неполный, наклонный краевидный киль (Van Jaarsveld, 1989), также называемый "краевой вершиной" (Roberts Reinecke, 1965). Этот признак широко распространен у Astroloba, Gasteria и Poellnitzia, в меньшей степени – у Haworthia, редко встречается у Aloe (A. variegata L. является известным исключением) и отсутствует у Chortolirion. Этот явление не считается гомологичным V-видному килю Kniphofia. Как и бугорчатость листьев, развитие очень острого киля часто рассматривается как защитная реакция от поедания травоядными животными.
6. Плотность соцветия
У видов Alooideae и Kniphofia соцветие – многоцветковый колос, либо простая или ветвистая кисть. У Alooideae соцветие развивается из пазухи листа, но фактически проявляется как верхушечное (Berger, 1908). Следовательно, стебли являются моноподиальными до формирования соцветия, после чего становятся симподиальными (Obermeyer, 1976). Таким образом, розетка, несущая соцветие, не отмирает после цветения, как это свойственно агавам. У большинства видов Alooideae и Kniphofia процесс цветения начинается от основания соцветия к его вершине (акропетальный). У некоторых центральноафриканских видов книфофии (G. W. Reynolds, 1966; Codd, 1968) и Aloe capitata Baker, однако, процесс цветения базипетальный (Verdoorn, 1970; Glen и Hardy, 1988).
У большинства видов книфофии, алоэ и ломатофиллума соцветия плотноцветковые. Это резко контрастирует с другими родами Alooideae, где соцветия - неизменно рыхлые кисти. Плотноцветковые кисти – вероятно, плезиоморфный признак для Alooideae, развившийся, по всей видимости, как приспособление к основному способу опыления. Hoffman (1988), например, убедительно показал, предлагая различное вознаграждение различным опылителям, что Aloe ferox использует дивергентные группы опылителей. На всей площади участков обитания последних, кисти алоэ бросаются в глаза яркими цветными заплатами на тусклом желтовато-сером фоне полуаридного южно-африканского пейзажа.
Хотя цветки Gasteria и Poellnitzia тоже ярко окрашены, их кисти не столь плотны, как у большинства видов Aloe и Kniphofia. Цветки видов Astroloba, Chortolirion и Haworthia обычно довольно неприметны и редко рассеяны на проволоковидной кисти. Это появляется как адаптация к специализированному синдрому опыления и, вероятно, является апоморфным состоянием.
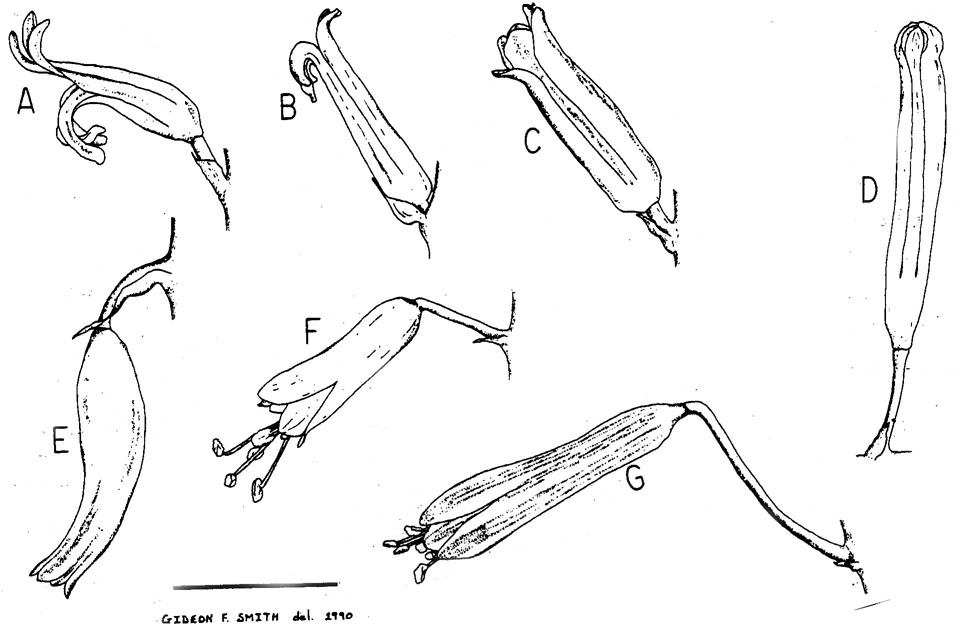
Рис. 1. Цветки представителей родов: A, Haworthia gracilis Poelln.; B, Chortolirion angolense (Baker) A. Berger; C, Astroloba foliosa (Haw.) Uitew.; D, Pellnitzia rubriflora (L. Bolus) Uitew.; E, Gasteria pulchra (Haw.) Haw.; F, Aloe tenuior Haw. var. rubriflora G. Reyn.; G, Lomatophyllum cf. purpureum (Lam.) T. Durand. (Все рисунки сделаны с живого материала. Масштаб = 5 мм.)
|
7. Форма цветка
Изменения структуры цветка различных родов Alooideae показаны на рис. 1. Даже при очевидности существования нескольких различных тенденций в развитии цветочной структуры у Alooideae (например, тенденция к зигоморфности у Chortolirion, Gasteria и Haworthia; см. также Rowley, 1967b, на Bulbine), цветки всех таксонов Alooideae и рода Kniphofia –трубчатые. Трубчатый околоцветник - превосходный резервуар для того изобилия нектара, что вырабатывают цветки большинства видов. Кроме того, высохшая трубка околоцветника способствует протеканию нектара от завязи к вершине цветка, где он скапливается в блестящую капельку. Это обычно совпадает с созреванием рыльца протандричных цветков и служит дополнительным атрактантом для потенциальных анемофильных опылителей (Mottram, 1977; Van Jaarsveld, 1987).
Околоцветник (или, правильнее, перигон) состоит из шести членов, каждые три из которых образуют мутовку. Эти шесть лепестков объединяются в короткую или заметную трубку, каковое объединение чаще отмечается в некоторых родах, таких как Gasteria. В тоже время, за исключением монотипичных родов Chortolirion и Poellnitzia, существует значительная внутриродовая изменчивость размеров и формы трубчатых цветков. Эта изменчивость была использована как основа для внутриродовой классификации родов Haworthia (Uitewaal, 1947b; Bayer, 1971) и Gasteria (Van Jaarsveld, 1989). Однако, Vaikos и др. (1978) показали, что, независимо от изменчивости морфологии цветка, у Aloe, Gasteria и Haworthia сосудистое снабжение элементов околоцветника удивительно единообразно. Трубчатые цветки родов Alooideae и Kniphofia – определенно синапоморфа для этих таксонов, что предполагает близкое филогенетическое родство между ними.
8. Срастание сегментов околоцветника
Срастание лепестковидных листочков околоцветника является вторым признаком, разделяющим Kniphofia и Alooideae и наводящим на мысль о монофилии. Однако, этот признак значительно изменчив даже в пределах одного рода. У некоторых видов Aloe, например, Aloe pictifolia Hardy (1976), сегменты околоцветника свободны в основании, тогда как у других таксонов слияние простирается почти до кончиков цветков (Aloe сер. Macrifoliae Haw.). Кроме того, другие виды Aloe (например, мадагаскарское A. albiflora Guillaumin) отличаются отсутствием отчетливой трубки околоцветника (Mathew, 1974). Хотя, в целом, морфология цветков Alooideae очень проста и довольно постоянна, возникновение множества отклоняющихся цветковых морф привело к опрометчивому выделению монотипных родов, особенно из Aloe (Aloinella Lemee не Cardot, Guillauminia A. Bertrand, Chamaealoe A. Berger). Сейчас, однако, все эти роды включены в Aloe.
9. Окраска цветков
Aloe, Gasteria, Lomatophyllum и Poellnitzia все имеют виды с ярко окрашеными цветками. В этих родах и в Kniphofia доминирует красная, желтая или оранжевая окраска цветков. Виды с двуцветными или даже трехцветными цветками также весьма обычны: Aloe excelsa A. Berger – красный и желтый (Leach, 1977); A. marlothii A. Berger – алый и зеленовато-кремовый (G. W. Reynolds, 1935); A. mutabilis Pill. – красный и зеленовато-желтый (G. W. Reynolds, 1950a); Gasteria acinacifolia – розовый и беловато-зеленый; G. nitida (Salm-Dyck) Haw. – красновато-розовый и желтый (Van Jaarsveld, 1989); Kniphofia sarmentosa (Andrews) Kunth – кораловый и лососевый (Codd, 1968). У некоторых видов Aloe внутривидовая изменчивость окраски цветков значительна; так, для A. chabaudii Schonl. были зарегистрированы желтые, оранжевые, розовые и красные формы (Leach, 1977). Как предложено для Gasteria (Van Jaarsveld, 1989), для Alooideae полностью монохромная окраска цветков – гипотетически наследуемый признак, однако, с точки зрения таксономии, он встречается ничтожно редко.
По сравнению с видами вышеупомянутых родов, окраска цветков представителей Astroloba, Chortolirion и Haworthia относительно тусклая; преобладает белый цвет, обычно с зеленым, коричневым или серым оттенком. Напротив, таксоны Aloe и Kniphofia с зелеными или белыми цветками редки, и виды типа A. albida (Stapf) G. Reyn., A. albiflora (Verdoorn, 1966), A. calcairophila G. Reyn. (G. W. Reynolds, 1961), A.chlorantha Lavranos (1973), A. compressa H. Perrier (Hardy, 1985) и A. prinslooi I. Verd. и Hardy (Verdoon, 1965) являются исключениями. Примечательно, что растения этих видов, как правило, относительно невысокие, как имеет место у Astroloba, Chortolirion и Haworthia, и цветки, по крайней мере, Aloe albida, A. compressa и A. prinslooi несут черты поверхностного сходства с двугубыми цветками Chortolirion и Haworthia. Виды Haworthia с ярко окрашенными цветками исключительно редки; лишь несколько клонов H. nortieri G. G. Smith имеют золотисто-желтые внутренние сегменты околоцветника (Anonymous, 1974), да некоторые экземпляры H. herbacea (Miller) Stearn sensu Bayer (1982) демонстрируют розовато-бежевые цветки. Гомологичность с розовыми и желтыми цветками некоторых видов Aloe еще не установлена.
По отношению к внешней группе Kniphofia, повсеместное наличие приглушенно-белых цветков у Astroloba, Chortolirion и Haworthia представляется унаследованным качеством.
10. Положение цветков
Представители трех родов с преобладающе беловатыми цветками, Astroloba, Chortolirion и Haworthia, несут распустившиеся цветки приподнятыми на вертикальных цветоножках (родовой признак Haworthia у Duval, 1809, читаем: " Calyx petaloideus, rectus, ... "). В этих родах угол между стебельком и цветоножкой всегда менее 90° (рис. 1). Ни цветки, ни семенные коробочки у растений этих таксонов не бывают висячими, что, вероятно, является наследственным признаком, развившимся в ответ на преобладающий специализированный энтомофильный синдром опыления.
Бутоны, отцветшие цветки и семенные коробочки некоторых видов Aloe, особенно Aloe серии Saponariae, часто несутся вертикально, но распустившиеся цветки отчетливо поникшие или распростертые. В случае большинства видов Alooideae, вертикальные семенные коробочки расположены на относительно высоких соцветиях, что предполагает адаптацию растений к рассеиванию семян ветром (анемохория). Это подтверждается тем фактом, что уплощенные треугольно-элиптические семена большинства видов имеют короткие (например, Haworthia spp.) или выступающие (например, Aloe variegata) бумаговидные крылышки. Однако, по крайней мере, на уровне видов, значение морфологии семян алоэ представляется таксономически несущественным (Kamstra, 1968).
Вертикальное (или висячее) положение распустившихся цветков не следует путать с односторонним расположением цветков, например, Aloe marlothii A. Berger, A. secundiflora Engler и Poellnitzia. Цветки последних таксонов расположены вертикально на более или менее горизонтальных осях соцветий. Такая диспозиция является адаптацией к опылению птицами – обычно крепкие горизонтальные ветви соцветий используются воздушными визитерами в качестве насестов. Так как цветки располагаются вертикально, опылители имеют свободный доступ к награде – нектару. Напротив, односторонне расположенные цветки некоторых видов Gasteria, например G. acinacifolia, имеющей плосковершинную метелку, всегда повислые. Поэтому, не зависающие воздушные опылители или приподнимают цветки своими хоботками, или, наклоняясь, достигают нектара.
11. Симметрия околоцветника
Как правило, зигоморфность редка в Asphodelaceae – симметрия цветков для Kniphofia и большинства родов Alooideae более-менее обычна (Rowley, 1967b; но см. Bayer, 1972). Зигоморфность цветков у Chortolirion, Gasteria и Haworthia восходит к исходному типу. Однако только в родах Chortolirion и Haworthia есть виды с двугубыми цветками. Что является устойчивой синапоморфой для этих родов. Они отличаются по цветкам от других родов Alooideae, у которых кончики долей околоцветника наклонно выступают до сильно загнутых. Примечательно, что у видов подрода Robustipeduncularis Uitew ex М. Bayer, основной группы в пределах Haworthia, цветки лишь слабо зигоморфны.
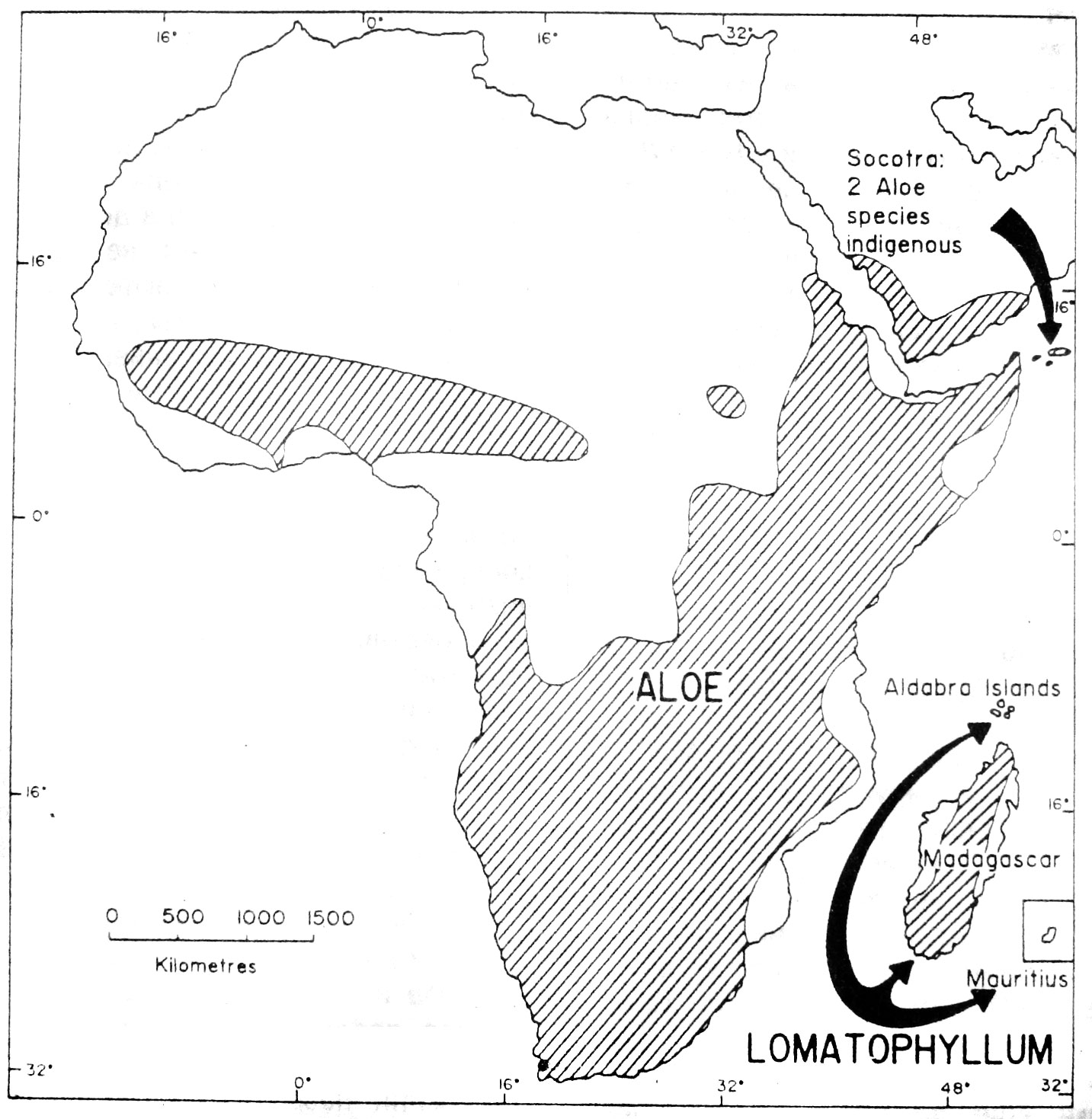
Рис. 3. Приблизительное географическое распространение родов Aloe и Lomatophyllum
|
Цветки представителей рода Gasteria (и, в меньшей степени, Poellnitzia) демонстрируют определенные тенденции к зигоморфности. Три наиболее заметных особенности цветков Gasteria: раздутое основание околоцветника, искривление конечной части трубки и сжатие ниже(?) шаровидной гастероидной части. Тем не менее, гастероидные цветки никогда не бывают двугубыми (рис. 1).
У большинства видов Aloe, Kniphofia и Lomatophyllum трубка околоцветника цилиндрическая, колокольчиковидная или воронкообразная. Aloe, однако, имеет множество видов с односторонне вздутыми (слабо зигоморфными) цветками (cf. Smith, 1990, на A. bowiea Schultes и Schultes f.). Очень немногие виды Aloe имеют явно зигоморфные цветки, известными исключениями являются A. albida и A. myriacantha (Haw.) Schultes и Schultes f., оба из секции Graminialoe. Однако, зевы цветков этих видов вздернутые, – признак, отсутствующий у Haworthia. Хотя истоки зигоморфности невполне ясны, это представляет продвинутое состояние, унаследованное от систематически плезиоморфного паттерна. Состояние сильно адаптивно, связано с большей селективностью на тип опылителя (Dahlgren и Rasmussen, 1983).
12. Цветочный аромат
Цветки подавляющего большинства видов Alooideae не имеют запаха. Цветочный аромат в подсемействе ограничен незначительным числом видов Аloe и Kniphofia. Все виды Kniphofia с ароматными цветками (K. brachystachya (A. Zahlbr.) Codd, K. parviflora Kunth, K. typhoides Codd, K. umbrina Codd) имеют короткие желтые или коричневатые цветки и образуют близко связанную группу (Codd, 1968).
В отличие от Kniphofia, виды Aloe с пахучими цветками не формируют такой группы, и запах отсутствует во многих таксонах, где этого можно было бы ожидать, например, у A. incospicua Plowes (1986). За исключением южно-африканского A. modesta G. Reyn., распространение пахучих алоэ ограничено Мадагаскаром (Bornman и Hardy, 1971; Van der Riet, 1977). Это подтверждает точку зрения Holland-а (1978), утверждавшего, что предки алоэ произошли из горной местности юго-восточной Африки еще до полного отделения Мадагаскара от Африки Мозамбикским проливом в середине мелового периода (100 миллионов лет назад) (Flores, 1970; McElhinny и др., 1976; но см. Croizat, 1968, на мадагаскарских видах Euphorbia). Это предположение подкрепляется тем фактом, что виды алоэ, встречающиеся на Мадагаскаре, не имеют двойников на Африканском материке и наоборот (G. W. Reynolds, 1955/1956, 1965), – фактом, свидетельствующем о древнем расхождении по отдельным эволюционным дорожкам. Мадагаскарские виды алоэ с пахучими цветками: A. compressa (Hardy, 1985), A. conifera H. Perrier (Hardy, 1989), A. cryptoflora G. Reyn., A. haworthioides Baker (Hardy, 1988) and A. suzannae (Popiel & Ellert, 1982). Цветки этих видов особенно сильно пахнут в вечерний период, привлекая ночных насекомых-опылителей (Hardy, 1985, 1989; но см. Glass и Foster, 1983). По отношению к внешней группе Kniphofia, отсутствие цветочного аромата у Alooideae отмечено, как унаследованное.
13. Положение пыльника
Большинство таксонов Alooideae имеет ярко оранжевые или желтые, спинкоприкрепленные, продолговатые или линейно-продолговатые пыльники с интрорзным раскрытием. В период пыления пыльники цветков большинства видов Kniphofia, Aloe и Lomatophyllum значительно высовываются из околоцветника. Напротив, у представителей Astroloba, Chortolirion, Gasteria, Haworthia и Poellnitzia пыльники втянуты в трубку околоцветника. Это различие довольно отчетливо и, несомненно, представляет значительный разрыв между Kniphofia плюс Aloe плюс Lomatophyllum и другими родами Alooideae. Предполагается, что цветки с втянутыми в трубку пыльниками только тогда могут быть эффективно опылены, когда потенциальный опылитель, активно протиснув ротовые органы (хоботок) в довольно узкую трубку околоцветника, достигает нектарной награды. В Alooideae предельное заключение пыльников в трубку найдено у Poellnitzia, где цветки вообще не открываются, за исключением трех узких щелей, образованных просветами между кончиками околоцветника. Этот признак аутапоморфный для рода. В случае цветков с выдвинутыми из околоцветника пыльниками, возможно опыление случайными посетителями, или посетителями, собирающими только пыльцу. По отношению к внешней группе, заключение пыльников в околоцветник выигрышно как апоморфа и, вероятно, представляет адаптацию специализированному синдрому опыления. Выдвинутые тычинки Aloe и Lomatophyllum могут, в конечном счете, оказаться полезной родовой синапоморфой в случае, если будущие исследования Asphodelaceae в целом докажут поверхностность сходства с Kniphofia.
14 и 15. Сахара нектара
Характерные цветки большинства таксонов Kniphofia и Alooideae нектароносны, и выделение нектара продолжается от одного до нескольких дней. Секреция нектара в подсемействе происходит посредством септальных нектарников (Schnepf и Pross, 1976). Размер цветков коррелирует с продуцированием нектара, крупноцветковые виды Aloe производят намного большее количество нектара, чем представители, например, родов Chortolirion и Haworthia (ср. Beyleveld, 1973, на выбранных видах Aloe серии Saponariae; и Mottram, 1977, Fox и Norwood Young, 1988, и Hoffman, 1988, на A. ferox).
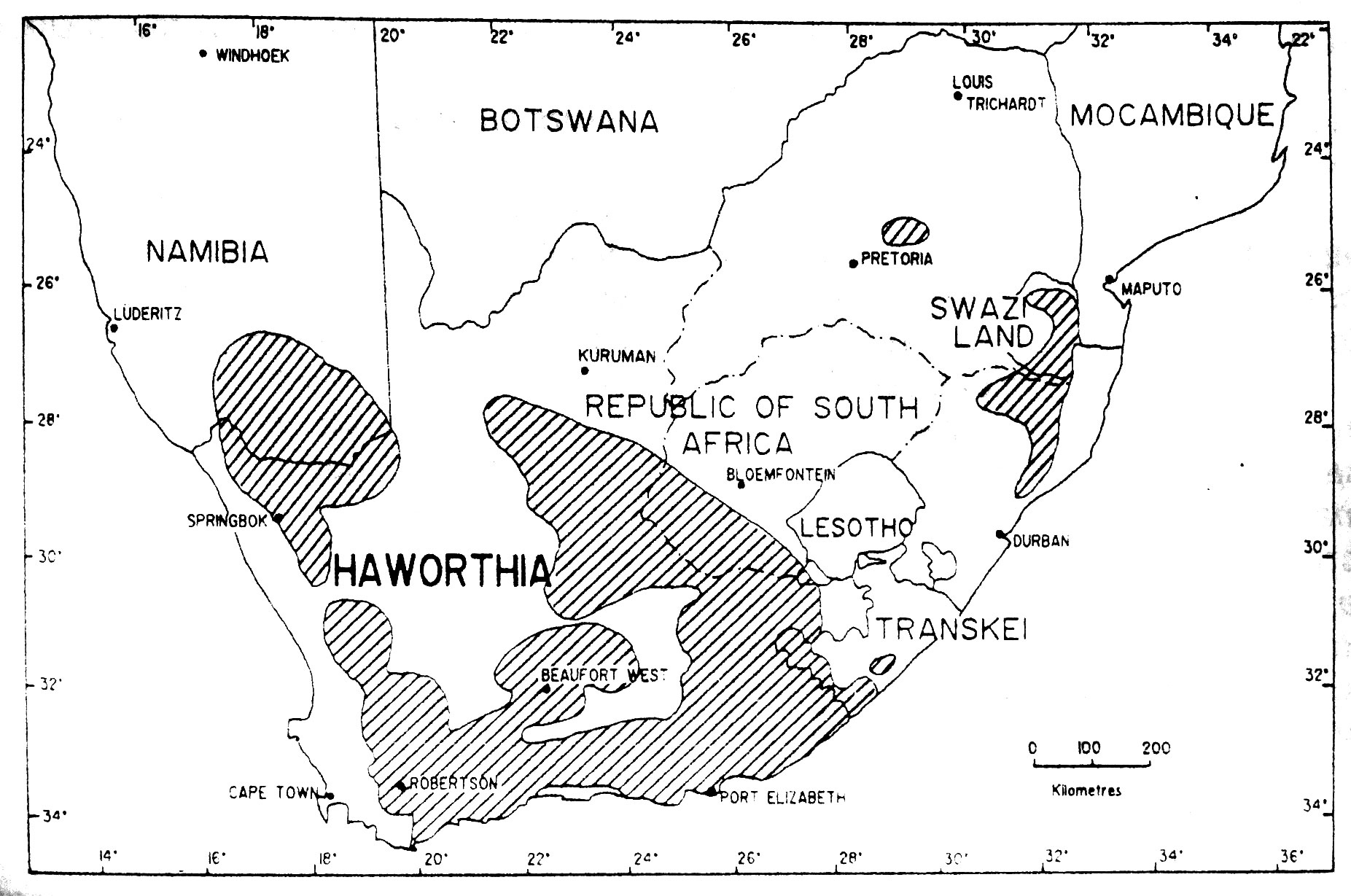
Рис. 4. Приблизительное географическое распространение рода Haworthia (Pilbeam, 1983; Bayer, 1986).
|
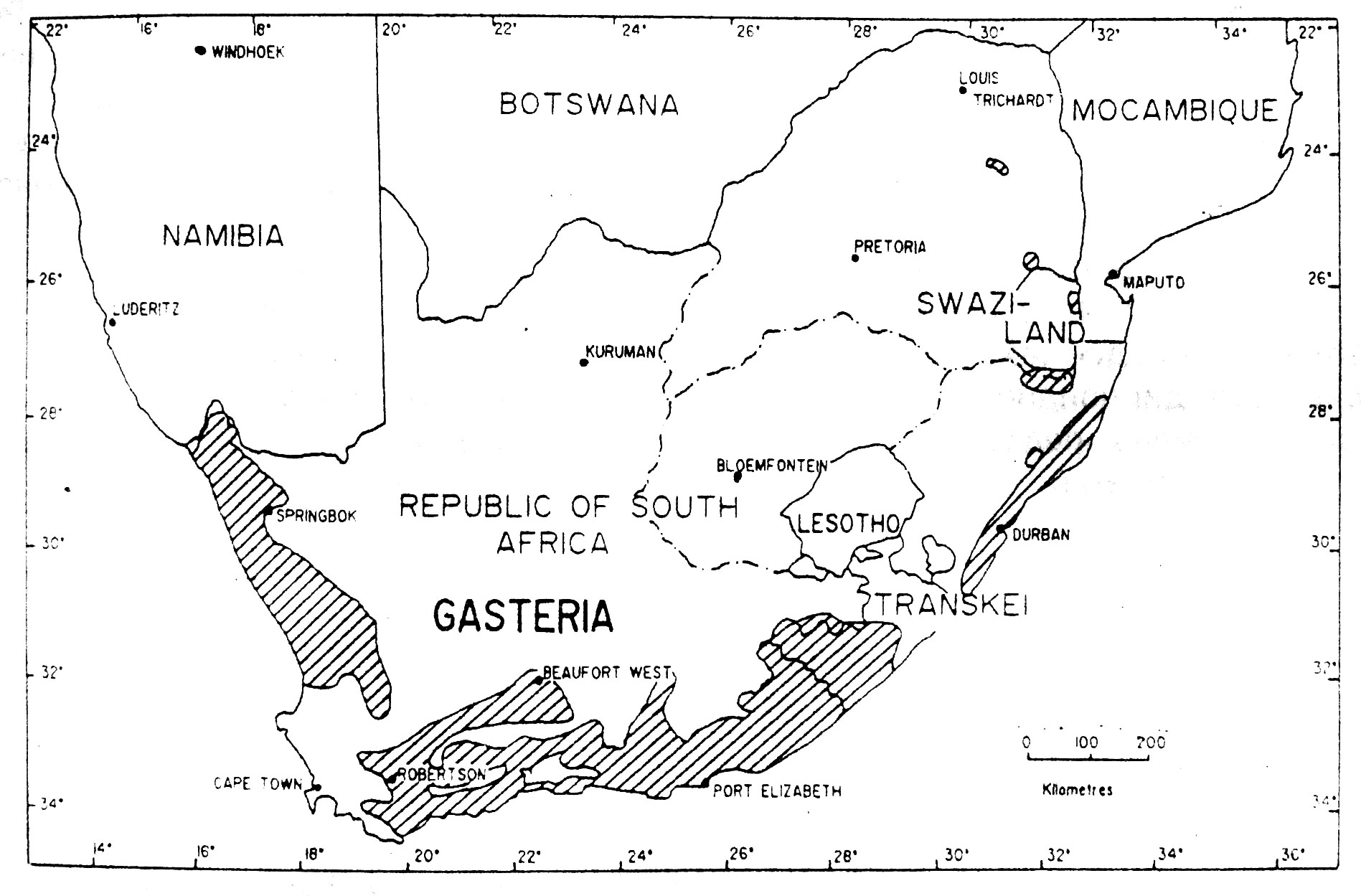
Рис. 5. Приблизительное географическое распространение Gasteria (Van Jaarrsveld, 1989).
|
Сахароидный состав нектара (глюкоза, фруктоза, сахароза) представительных образцов Kniphofia и всех родов Alooideae был недавно определен. Это исследование показало, что сахароидный состав в пределах рода замечательно постоянен. Были выделены три типа нектара: алоидный тип, представленный Aloe (включая Chamaealoe), Kniphofia, Lomatophyllum и Poellnitzia; гастериоидный тип – представлен только Gasteria; и хавортиоидный тип – Astroloba, Chortolirion и Haworthia. Алоидный тип отличается тем, что сахароза фактически отсутствует; фруктоза же и глюкоза представлены в более-менее равном количестве. Для гастериоидного типа характерно преобладание сахарозы; фруктозы и глюкозы примерно поровну. В хавортиоидном типе сахароза также доминирует, но глюкозы много больше, чем фруктозы. По отношению к внешней группе, преобладание сахарозы (признак 14) и ассиметричная пропорция фруктозы и глюкозы (признак 15) рассматриваются как апоморфные.
16. Основное число хромосом
Большинство видов Alooideae цитологически изученны. Эти исследования начались в начале ХХ столетия (ср. Taylor, 1925), в результате чего имеется обширная библиография по цитологии этой группы (для обозрения см. Muller, 1941; Riley, 1959a,b,c; Brandham, 1971, 1983; Riley и Majumbar, 1979). Alooideae – одна из наиболее однородных групп в отношении как числа хромосом, так и выраженно бимодального кариотипа. Все виды имеют одно и тоже основное число хромосом (х = 7), с 4 длинными и 3 короткими хромосомами. В подсемействе в целом отклонение от основного диплоидного кариотипа (2n = 14) встречается очень редко (Brandham, 1969). Данный признак является синапоморфой для всех таксонов Alooideae. В пределах Alooideae существует множество внутри- и межродовых гибридов (Rowley, 1982). Это ясно свидетельствует о цитогенетически тесных родственных отношениях, которые существуют между видами подсемейства (Rollins, 1953).
Базируясь на общем размере генома и увеличении бимодальности, Brandham (1983) показал, что, по крайней мере у алоэ, существует градация от наименьших хромосом у видов, сохраняющих набор презиоморфных признаков (A. tenior: актиноморфные цветки; слабые вьющиеся стебли; мезофит), до наибольших – у видов с морфологическими апоморфами (A. peckii Bally & I. Verd.: бесстебельное; чрезвычайный ксерофит).
В отличие от Alooideae, Kniphofia имеет основной набор из 6 хромосом (2n= 12). Это представляет четкий разрыв между этими двумя таксонами и является определенным барьером для обмена генами (De Wet, 1960). По отношению к внешней группе Kniphofia, основной хромосомный набор х = 7 является выигрышным, как апоморфа.
Филогенетические подтексты
Без опыта в кладистике, исследования Alooideae были сделаны поспешно, и результаты фенетических исследований неубедительны (для сравнения см. наше введение). На основе отличий кариотипа и характерной морфологии листа предлагается считать подсемейство Alooideae монофилитичным. В этом отношении положение рода Bulbine (подсем. Asphodeloideae) предстает проблематичным, поскольку некоторые из его видов имеют кариотипы и морфологию, подобные таковым некоторых таксонов Alooideae (ср. Spies и Hardy, 1983, на B. latifolia (L. f.) Schultes и Schultes f.; Rowley, 1954, гл. обр. на Bulbine). Наше исключение Bulbine из Alooideae основано на нетрубчатых и обычно однородно желтых цветках (Trager, 1984), свободных листочках околоцветника, и недостатке производства нектара, предполагая, что сходство с Alooideae негомологично.
В отличие от Alooideae, у Kniphofia отсутствует суккулентность листьев, контуры листа в поперечном сечении V-образные (а не серповидные или ладьевидные), и основное число хромосом х = 6, а не х = 7. Мы расцениваем трубчатость цветков и срастание сегментов околоцветника как достаточные свидетельства оправданности выбора Kniphofia как внешней группы по отношению к Alooideae. Род Kniphofia был отделен от Alooideae на основе, помимо прочего, анатомических различий в строении листьев (Baijnath, 1980; Dahlgren и др., 1985). Типичные сосудистые пучки таксонов Alooideae sensu Dahlgren и др (1985) имеют хорошо развитую чашевидную структуру тонкостенных клеток паренхимы во флоэмном полюсе. Их часто упоминают как алоидные клетки. Напротив, Kniphofia имеет хорошо обозначенные волокна, присутствующие в чашевидной структуре и в ксилемном, и во флоэмном полюсах (Beaumont и др., 1985). Присутствие алоидных клеток значительно меняется в пределах между таксонами (а иногда даже в пределах листа отдельного представителя) Alooideae, и этот признак нелегко поддается филогенетической интерпретации. Однако, заключение Beaumont и др. (1985), что паренхимное состояние (преобладание тонкостенных клеток по сравнению со склеренхимой) наследуемо, подтверждает наш выбор Kniphofia в качестве внешней группы по отношению к Alooideae.
На кладограмме (рис. 2) Aloe и Lomatophyllum показаны, как лежащие ближе к основанию всего подсемейства, чем остальные рода. Напротив, Astroloba, Chortolirion и Haworthia наиболее далеко продвинуты по отношению к общим предковым формам, а Gasteria и Poellnitzia занимают промежуточное положение. Линия Alooideae определена четырьмя апоморфами, одна из которых (стеблевой характер произрастания) имеет изменение выше (Chortolirion). Противоречие, которое окружило родовое определение границ, у Alooideae отражено недостатком автоапоморф для некоторые из родов, поддержанных в этой работе. Только Gasteria (три отчетливые апоморфы), Chortolirion, Lomatophyllum и Poellnitzia (одна отчетливая апоморфа для каждого) действительно хорошо определены. Однако, примечательно, что три последних рода, Chortolirion, Lomatophyllum и Poellnitzia, печально известны таксономической путаницей. В прошлом их то объединяли с Aloe, Astroloba и Haworthia, то вновь отделяли. Возвращение к концепции Duvalian алоидной классификации, в которой только три "суперрода" – Aloe, Gasteria и Haworthia, несовместимо с нашим результатом. Включение Astroloba и Chortolirion в Haworthia, однако, снабдило бы род по крайней мере двумя апоморфами (признаки 11 и 15) и улучшило бы определение родовые границ в пределах подсемейства. Нет никаких убедительных свидетельств монофилитичности, в строгом смысле, Haworthia, и возможное расширение родовой концепции кажется неизбежным. Точно так же, существующая родовая концепция Aloe открыта для спекуляций. Наш результат выдвигает на первый план главную слабость существующей системы – что Aloe и Haworthia могут и не быть монофилитичными. Будущие исследования должны быть направлены на решение этой основной проблемы.
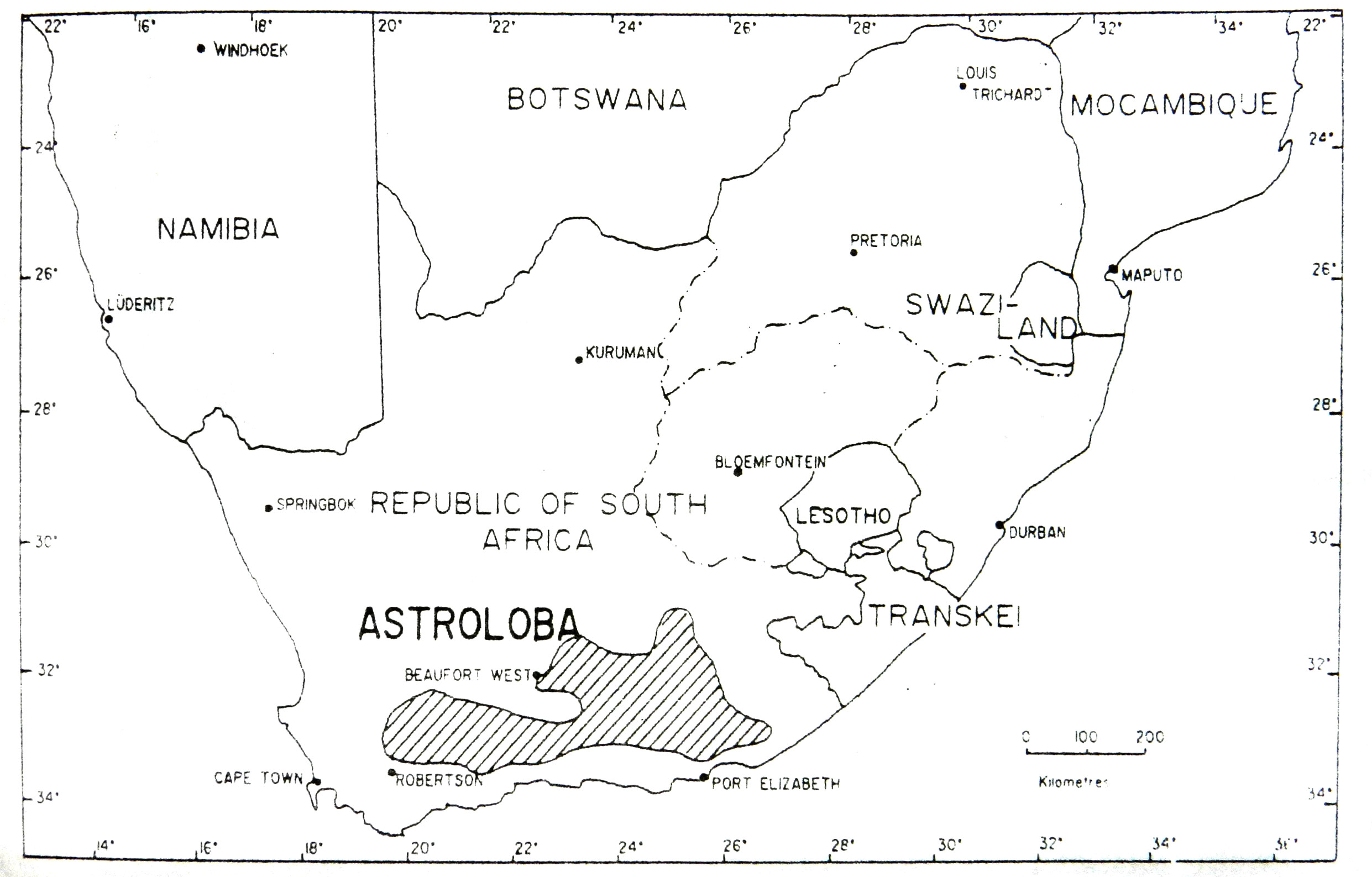
Рис. 6. Приблизительные границы ареала рода Astroloba ( Roberts Reinecke, 1965).
|
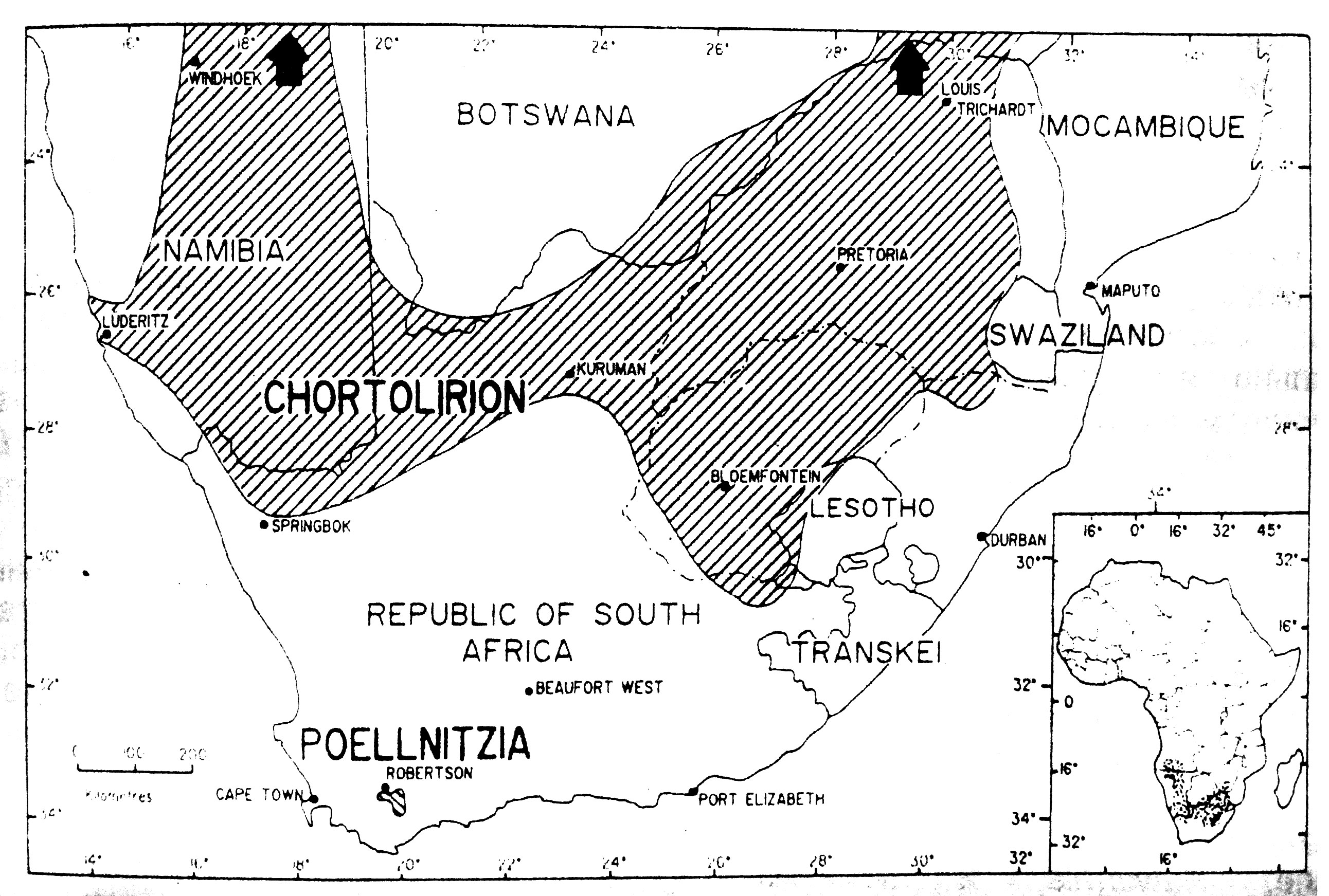
Рис. 7. Приблизительные границы распространения родов Chortolirion и Poellnitzia. Вполне вероятно, что Chortolirion распространён дальше на север в Ботсвану, и дальше на юг в Наталь. Когда он не цветёт, его легко не заметить (Smith, 1985).
|
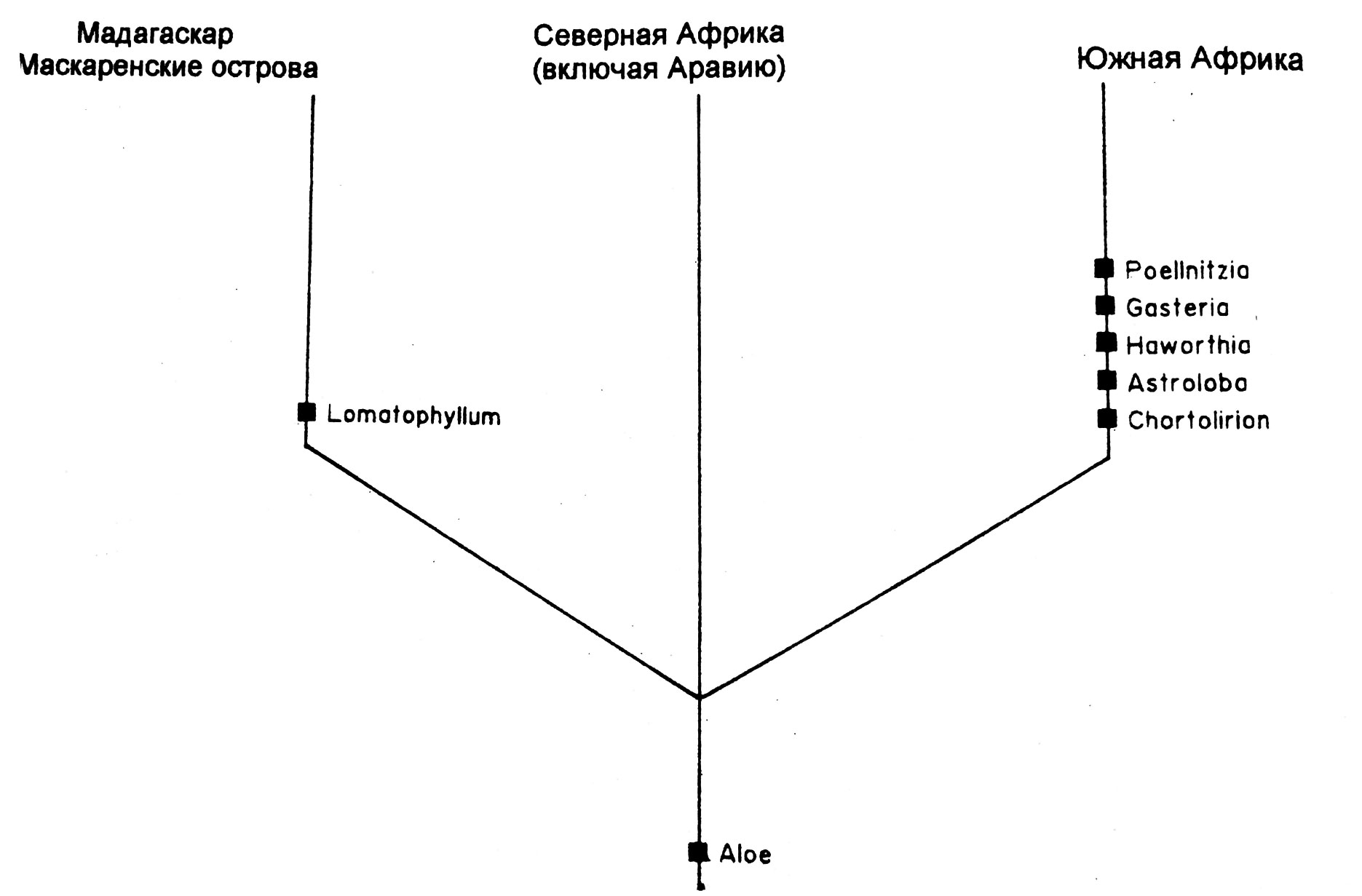
Рис. 8. Территориальная кладограмма подсемейства Alooideae, показывающая три основных территории. Филогения родов на Рис. 2 не даёт информации о таких двух важнейших событиях, как отделение от Африки Мадагаскара и одновременная изоляция Южной Африки от остального континента.
|
Географические соображения
Приблизительно географическое распространение родов Aloe, Astroloba, Chortolirion, Gasteria, Haworthia, Lomatophyllum и Poellnitzia показано на рис.3-7. Aloe встречается в Африке южнее Сахары, на Аравийском полуострове, Мадагаскаре и Сокотре. Распространение Kniphofia также является, главным образом, палеотропическим (Судано-ангольский и Карру-Намибский регионы Африканского подцарства Палеотропического царства, sensu Takhtajan, 1969) и, в значительной степени, совпадает с ареалом Aloe, однако род отсутствует на Сокотре, в Намибии и Ботсване (Codd, 1968). Виды Kniphofia склонны, похоже, к влажным биотопам и найдены, в основном, вдоль горных цепей. По контрасту, виды Aloe экологически гетерогенны в пределах ареала рода и разнообразно представлены в почти каждой возможной среде обитания, включая пустыни, степи, саванны и сравнительно влажные прибрежные леса. С точки зрения количества видов, и Aloe, и Kniphofia сконцентрированы теперь в Южной Африке. Распространение мясистоплодного Lomatophyllum ограничено несколькими из Маскаренских островов у юго-восточного побережья Африки.
Ареалы Astroloba, Gasteria и Haworthia довольно похожи. Эти три рода являются эндемиками Южной Африки и более-менее ограничены полуаридными прибрежными районами с сухим летом, ниже внутреннего уступа субконтинента. Gasteria и Haworthia имеют заходы в аридные речные долины Наталя, Свазиленда и восточного Трансвааля, с единственным таксоном Haworthia – H. venosa subsp. tessellata (Haw). М. Bayer, встречающейся в климатически суровой центральной части южной Африки. Распространение Astroloba более ограничено, чем Gasteria и Haworthia, обычно их находят в несколько более засушливых местах обитания финбоша и в биомах суккулентного Карру Южной Африки. Эти три рода, как и Aloe имеют сравнительно большое число местных видов и эндемиков аридных субтропических переходных зарослей восточной части Капской провинции, где они проявляют признаки активной специализации (см. Court, 1981; Smith и Marx, 1990; и там же ссылки).
Из всех родов Alooideae, монофилетическая Poellnitzia имеет наименьший ареал. Этот род найден только в районах Робертсон и Боннивале на юго-западе Капской провинции. По контрасту, другой монофилитический род Chortolirion широко распространен в южноафриканских степях с дождливым летом. Род, однако, отсутствует в районе с дождливой зимой в южной Намибии.
Ареал Alooideae включает три основных компонента: Африку, Мадагаскар с прилегающими островами и южную Аравию. Как показано на рис. 8, имеется некоторое свидетельство ввикарирования в фитогеографической истории подсемейства. Два главных события обозначены, даже при том, что эффекты, как кажется, были частично затенены последующими событиями: (1) отделение Мадагаскара от Африканского материка (средний Мел, около 100 миллионов лет назад; G.Flores, 1970, и McElhinny & al., 1976); (2) разделение Южной Африки и остальной части Африки, возможно в результате аридификации, которая началась в течение Олигоцена (около 38-26 миллионов лет назад) и продолжилась в Миоцене и Плиоцене (около 26-7 и 7-2 миллиона лет назад, соответственно) (Goldblatt, 1978; Deacon, 1983; Raven, 1983; Van Jaarsveld, 1987). Современное распространение родов неинформативно относительно последователь-ности этих событий, то есть, произошло ли отделение Южной Африки до или после отделения Мадагаскара.
Заключение
По имеющимся данным можно выстроить лишь одну ясную филогению родов Alooideae. Как ясно видно из наших результатов, если включение Lomatophyllum в Aloe и включение Chortolirion и Astroloba в Haworthia может быть оправдано, то относительно родового статуса Poellnitzia могут быть некоторые сомнения. Мы предлагаем, однако, сохранить родовые концепции, используемые в этой работе, до тех пор, пока детальные таксономические ревизии (особенно родов Aloe, Haworthia и Lomatophyllum) не станут доступными.
Точно так же, перемещение Kniphofia в Alooideae кажется преждевременным на этой стадии, т. к. только анализ семейства в целом мог бы продемонстрировать что такое перемещение - наименее аргументированное решение. Широко концептуальный подход данной работы дает, однако, некоторую ясность в инфраподсемейной филогении Alooideae.
Следующий ключ должен помочь различать Kniphofia (внешний род) и роды Alooideae, признанные в этой работе:
| 1. Листья травянистые, мягкие, несуккулентные, непятнистые, не имеющие отчетливых колючек, в прикорневых розетках (апикальных у Kniphofia northiae Baker)
| Kniphofia
|
| – Листья толстые, твердые, суккулентные, пятнистые, часто окаймленные колючими зубцами, в прикорневых или апикальных розетках, либо стеблевые
| 2
|
| 2. Плод ягода
| Lomatophyllum
|
| – Плод коробочка
| 3
|
| 3. Коробочка апикально заостренная, в основании вздутая, цветки обычно < 15 мм длиной, зигоморфные
| Chortolirion
|
| – Коробочка апикально закругленная или тупая, в основании ризоматозная (если изредка выпуклая, то цветки > 15 мм длиной, актиноморфные)
| 4
|
| 4. Сегменты околоцветника апикально сходящиеся
| Poellnitzia
|
| – Сегменты околоцветника апикально расходящиеся или отогнутые
| 5
|
| 5. Цветки при расцветании повисают, трубка околоцветника изогнута вверх
| Gasteria
|
| – Цветки при расцветании не повисают, трубка околоцветника прямая или изогнутая вниз
| 6
|
| 6. Околоцветник двугубый, < 15 мм длиной, зев не вздернутый
| Haworthia
|
| – Околоцветник правильный (если изредка слабо двугубый, то цветки > 15 мм длиной, зев вздернутый)
| 7
|
| 7. Цветки обычно ярко окрашенные, мясистые, тычинки длиной с околоцветник или длиннее
| Aloe
|
| – Цветки тускло окрашенные, тонкие, тычинки заключены в околоцветник
| Astroloba
|






