ЖУРНАЛ "КАКТУСЫ И ДРУГИЕ СУХОЛЮБИВЫЕ РАСТЕНИЯ" - выпуск 15-16 (1-2 за 2003 год)
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ СУХОЛЮБИВЫХ РАСТЕНИЙ
Часть 1. Введение. ВЫХОД РАСТЕНИЙ ИЗ ВОДНОЙ СРЕДЫ В АТМОСФЕРУ
(Изложение доклада, прочитанного на собрании РОИКС в марте 2003 г)
Синёв И.Е.
Возникновение жизни на Земле - одна из самых интересных и загадочных проблем, имеющая важнейшее не только естественно-научное, но и философское значение. Живых свидетелей процессов зарождения биологической жизни на нашей планете нет. А современные условия исключают возможность повторения этих процессов.
Тем не менее существует каменная летопись Земли – ископаемые остатки живых организмов, и ископаемые свидетельства не биологических процессов, имевших место на нашей планете в разные периоды её существования. Изучение их геологами, палеонтологами, палеоботаниками и многими другими специалистами смежных специальностей позволяет нам в общих чертах представить себе историю зарождения и эволюции жизни на Земле.
В настоящее время считается общепризнанным, что жизнь зародилась приблизительно 3.5 миллиардов лет назад.
Экологические условия на Земле 3,5 млрд. лет назад.
Сумма накопленных знаний позволяет предположить, что условия для зарождения жизни были достаточно благоприятными. Атмосфера первобытной Земли была перенасыщена углеродом – химической основой органического вещества, и водяным паром, обладала пониженной прозрачностью и содержала другие химические вещества, необходимые для зарождения жизни, в частности, азот. По данным химического анализа газовых пузырьков в несколько более поздних породах - кварцитах курумканской свиты Алданского щита (Салоп Л. И. по: Ушаков С. А., Ясаманов Н. А., 1984), в этих пузырьках содержится около 60% углекислого газа и около 35% H2S, SO2, NH3, HCl, HF. Высокое содержание СО2 в атмосфере обусловливало резко выраженный парниковый эффект. Температура поверхности Земли, оценки которой проводились по породам с возрастом от 2,3 до 1,3 млрд. лет, была в диапазоне 40-70°С. Соответственно, температура верхних слоёв воды также была повышенной, сравнительно с современными условиями, но не превышала, по-видимому, 40-45°С. Широтная зональность климата отсутствовала, температурный, но не световой режим на полюсах мало отличался от такового в экваториальной зоне. Интересно в связи с этим, что атмосфера ближайшей к нам планеты Сонечной системы Венеры, по данным инструментальных измерений со спущенных на её поверхность исследовательских зондов, также состоит главным образом из углекислого газа.
Углерод, как известно, “склонен” к образованию молекулярных цепочек. Не случайно химии известны лишь несколько тысяч неорганических (без атомов углерода) химических соединений, и почти в тысячу раз больше – органических. В ходе экспериментов, бывших особенно популярными в середине прошлого века, было доказано, что вполне возможно абиотическое возникновение таких органических молекул, как альдегиды, нитриты, аминокислоты (глицин, аланин, аспарагиновая и глутаминовая кислоты), моносахариды, пурины, порфирины нуклеотиды и т. п. От этих и массы других “биомономеров” до первых клеточных микроорганизмов дистанция громадной протяжённости. Но в конечном итоге она была пройдена. И микроорганизмы заселили первобытные водоёмы: реки, озёра и океан.
Самые ранние следы предположительно биогенного происхождения найдены в строматолитах Австралии – осадочных структурах, имеющих возраст около 3,5 млрд. лет. Здесь же в Австралии в породах с возрастом не менее 2,8 млрд. лет были найдены следы микробиологических сообществ.
Важнейшим революционным этапом в развитии органической жизни на нашей планете следует считать появление фотосинтезирующих микроорганизмов примерно 2200-2300 млн. лет назад. Достоверно установлено, что сине-зелёные водоросли появились более 2 млрд. лет назад. В кремнистых сланцах Северной Америки (формация Ганфлинт) была найдена ископаемая микрофлора, свидетельствующая о существовании свыше 2 млрд. лет назад клеток нескольких типов. Мощные горизонты строматолитовых известняков и доломитов на всех континентах, образовавшихся 2,2-2,3 млрд. лет назад в результате жизнедеятельности сине-зелёных водорослей и бактерий, свидетельствуют, что к этому времени живые организмы не только обитали во всех водоёмах Земли, но и заметно воздействовали на окружающую среду. В первую очередь это воздействие отразилось на химическом составе атмосферы: именно в это время - примерно 2 млрд. лет назад, бескислородные до этого гидросфера и атмосфера стали малокислородными – появился свободный кислород, хотя и в количествах, не превышающих несколько десятых долей процента. Древнейшие окисленные осадочные породы – красноцветы – имеют возраст несколько менее 2 млрд. лет. Дальнейшее повышение содержания свободного кислорода в гидросфере привело к увеличению размеров клеток ископаемых микроорганизмов примерно 1,5 млрд лет назад (Соколов Б. С. - Органический мир Земли на пути к фанерозойской дифференциации. М.: “Наука”, 1975, 20 с.).
Следующим этапным шагом эволюционного развития жизни на Земле стал расцвет водных многоклеточных организмов в Позднем Протерозое – около 800-700 млн. лет назад. В Венде уже существовали крупные животные (бесскелетные кишечнополостные и черви) и растительные организмы, оставившие многочисленные ископаемые следы.
Таблица 1. ПЕРИОДИЗАЦИЯ ГЕОЛОГИЧЕСКОЙ ИСТОРИИ ЗЕМЛИ
| ЭРА | ПЕРИОДЫ | ХРОНОЛОГИЯ
(млн. лет) | РАЗВИТИЕ РАСТИТЕЛЬНОГО МИРА
|
| Доархейская | | 4500 - 3500 |
|
| Архей | | 3500 - 2600 | Возникновение протобионтов
|
| Протерозой | Ранний и Средний | 2600 - 800 | Возникновение клеточных организмов, появление растительных фотосинтезирующих организмов – одноклеточных водорослей.
Возникновение многоклеточных организмов.
|
| | Венд | 800 - 570 | Расцвет многоклеточных организмов, в том числе водорослей.
|
| Палеозой | Кембрий | 570 - 500 | Дивергентная эволюция водорослей
|
| | Ордовик | 500 - 440 | Большое разнообразие водорослей
Выход водорослей в атмосферу
|
| | Силур | 440 – 405 | Появление наземных растений-амфибий – риний и псилофитов.
|
| | Девон | 405 – 350 | Возникновение плауновидных, хвощевидных, папортниковидных, голосеменных, грибов
|
| Мезозой | Карбон | 350 – 285 | Появление первых хвойных
|
| | Пермь | 285 – 230 | Вымирание растений-амфибий
|
| | Триас | 230 – 195 | Появление зональности климата
|
| | Юра | 195 – 137 | Папоротники и голосеменные.
Первые засухоустойчивые растения.
|
| | Мел | 137 – 67 | Расцвет покрытосеменных растений Появление ксерофитов
|
| Кайнозой | Палеоген | 67 - 54 | Господство покрытосеменных,
расцвет флоры ксерофитов.
|
| | Эоцен | 54 - 37 | Появление первых суккулентов
|
| | Олигоцен | 37 – 26 |
|
| | Миоцен | 26 – 8 |
|
| | Плиоцен | 8 – 2 |
|
| | Плейстоцен | 2 - наст. вр. |
|
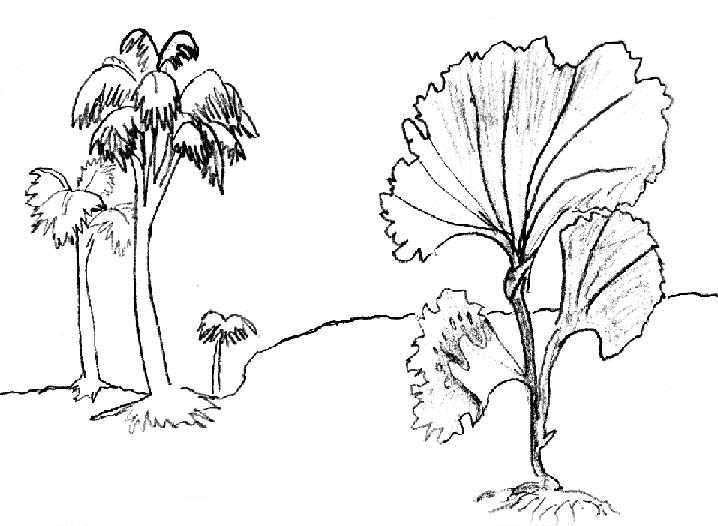
Рис. 1. Членистое строение слоевищ некоторых современных водорослей (слева Лессония) |
Главным “геологическим” результатом жизнедеятельности водных организмов, помимо выделения свободного кислорода, было связывание в органическом веществе и фоссилизация громадных количеств углерода, ставшего впоследствии ископаемым топливом.
Экологические условия 500 млн. лет назад. Изучение довольно скудной информации относительно природных условий, имевших место в Ордовике и Силуре (500-405 млн. лет до нашей эры) позволяет сделать предположение о том, что атмосфера Земли по-прежнему была перенасыщена углекислым газом. За 3 млрд. лет в результате жизнедеятельности водных растений его концентрация уменьшилась в несколько раз и появился свободный кислород, количество которого в атмосфере в рассматриваемый период не превышало нескольких процентов. Перенасыщенность атмосферы углекислым газом обусловливала резко выраженный парниковый эффект, низкую прозрачность воздуха и, соответственно, значительно пониженную, по сравнению с современными условиями, освещённость в водной среде. Климатическая зональность отсутствовала.
Мы не знаем, что происходило в ту далёкую эпоху 500 миллионов лет назад. Тем не менее имеющиеся данные позволяют предположить, что адаптация к воздушной среде пол-миллиарда лет назад могла происходить одновременно у водорослей из нескольких отделов, в первую очередь имевших элементы разделения слоевища на отдельные ткани, а также членисто-мутовчатое строение слоевищ (Рис. 1), но также и из других. Представляется сомнительным выводить все наземные растения от одного общего предка. Возможно, какие-то зелёные водоросли-гаметофиты, не имевшие членистого строения слоевищ, могли дать начало предкам печёночных мхов. Другие водоросли, спорофиты, возможно, из вымершего к настоящему времени отдела, или подотдела, например, бурых водорослей (целлюлоза современных бурых водорослей существенно отличается от целлюлозы высших растений), могли стать предками первых атмосферных растений.
Выход из условий невесомости в водной среде возможен, на наш взгляд, по трём моделям:
- “выползание”, когда водоросли, покрывающие дно, стелятся по поверхности окружающей водоём суши;
- “подъём”, когда водоросль уже имеет вертикальный подводный побег, способный принять на себя вес верхней, “атмосферной”, части побега.
- можно также предположить более широкое распространение в прошлом водорослей типа саргасовых, плавающих у поверхности воды за счёт воздушных полостей, играющих роль поплавков, и также имеющих возможность ассимилировать углекислый газ непосредственно из атмосферы. Однако прогрессирующее уменьшение насыщенности атмосферы углекислым газом могло прервать эволюцию в этом направлении.
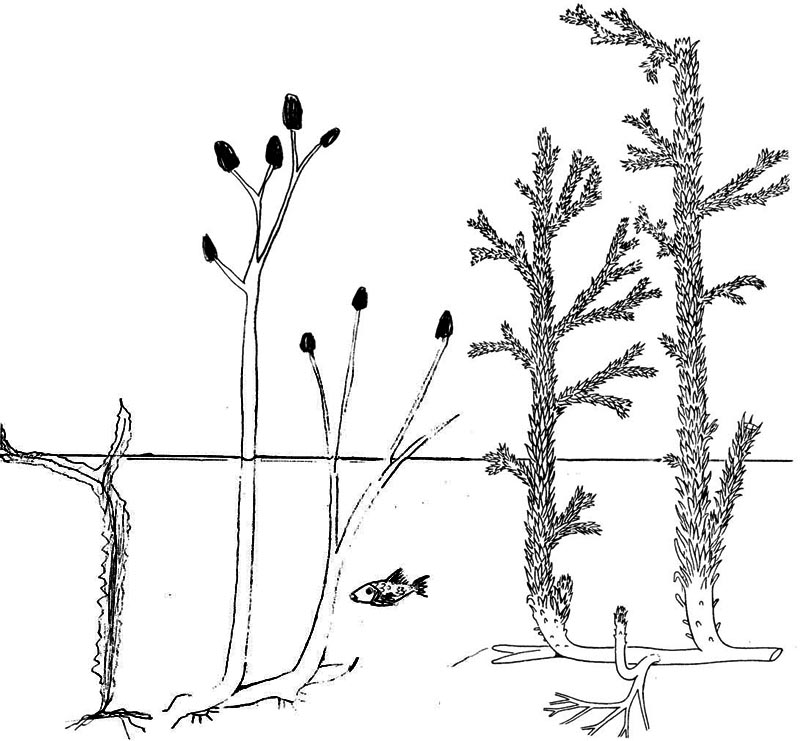
Рис. 2. Первые “наземные” растения, слева направо: риниофиты Тениокрада и Риния большая, справа ископаемое плауновидное Астероксилон, обитавшие примерно 400 млн. лет назад. |
Конкурентная борьба за свет в сообществах донных растений пресноводных водоёмов могла вполне стать причиной отбора и эволюции в направлении большей механической прочности базальной части слоевища у каких-то водорослей, что позволило им выносить верхние части слоевища к поверхности воды. Нельзя исключить и то, что стимулом к усилению механической прочности базальной части слоевища мог стать отбор на надводный многогнездный спорангий. Это явление отмечено для представителей рода Тениокрада (Taeniocrada), виды которых образовывали подводные заросли с поднимавшимися над водой спорангиями (Рис. 2). Как бы то ни было, но любая часть водного растения, оказавшись в атмосфере, в среде с плотностью в 800 раз меньшей, чем вода, тут же приобретала вес, который ложился на нижние части растения. Именно отсутствие опоры не позволяло плавающим водорослям конкурировать за свет выше уровня поверхности воды. Жёсткость базальной части слоевищ в условиях снижения уровня поверхности воды приводила к тому, что верхние части растений оказывались в атмосфере и в процессе эволюции в конечном итоге приспособились к воздушной среде.
Этому благоприятствовали несколько факторов:
- перенасыщенность атмосферы углекислым газом - на момент выхода растений в атмосферу содержание СО2 в воздухе составляло порядка 30% - то есть примерно в тысячу раз больше, чем в настоящее время (0,03%), что не могло не стимулировать развитие у некоторых водорослей способности к поглощению углекислого газа непосредственно из атмосферы;
- меньший, чем в настоящее время, уровень освещения земной поверхности, в частности, отсутствие прямой солнечной радиации
- максимально высокая влажность воздуха над водной поверхностью, близкая к 100%, что увеличивает жизненный потенциал водных растений в воздушной среде.
Не следует воспринимать буквально выражение “выход растений на сушу”. Невозможно представить. чтобы такой процесс произошёл скачкообразно. Для того, чтобы атмосферная часть растения не высыхала, у предков высших растений должны были выработаться органы и ткани, отсутствовавшие, или же имевшиеся лишь в зачаточном состоянии у водорослей:
- эпидерма – представляет собой слой поверхностных клеток, формирующий покровную ткань. Эпидерма предохраняет растения от высыхания, а также от проникновения внутрь микроорганизмов. Эпидерма возникла в результате соответствующей специализации наружных клеток растений. Чем более приспособлено растение к наземным условиям, тем сильнее развита эпидерма.
- устьица - слой эпидермы защищает ткани растения от высыхания в атмосфере, но одновременно препятствует газообмену и лишает, таким образом, смысла адаптацию к атмосферным условиям. Поэтому устьица развивались одновременно с эпидермой. Через устьица проходит интенсивная диффузия водяного пара, углекислого газа и кислорода. Каждое устьице состоит из пары замыкающих клеток и устьичной щели, которая представляет собой межклетник, который ведёт в межклеточное пространство, называемое дыхательной полостью. Дыхательные полости пронизывают ткань фотосинтезирующего органа, обеспечивая газообмен с воздушной средой.
- проводящая система - предназначена для снабжения водой атмосферной фотосинтезирующей части растения, которая в результате газообмена с атмосферой непрерывно испаряет водяной пар.
Возникновение эпидермы, устьиц и проводящей системы позволило растениям “выйти на сушу”.
В итоге в отложениях с возрастом свыше 415 миллионов лет были найдены первые известные нам растения риниофиты (Тахтаджян А.Л., Жизнь растений, т. 4, 1978). Благодаря простоте своего строения, риниофиты больше были похожи на водоросли, чем на высшие растения. Они характеризовались верхушечным расположением спорангиев, слабо развитой проводящей системой, примитивным анатомическим строением, отсутствием листьев и корней.
В течение многих десятков миллионов лет после выхода в атмосферу растения заселяли не собственно сушу, а водоёмы на материках: русла рек, озёра, лагуны и т.д. Фактически это были растения-амфибии. Характерной их особенностью была тесная связь с водной средой.
Реальный выход растений на сушу произошёл несколько позднее, но об этом в следующем номере.
(продолжение в следующем номере)






