| Список статей | Контакт с автором | English | RUS=Win1251 |
МОРФО-ФИЗИОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ КАЛЛУСНЫХ КУЛЬТУР ПШЕНИЦЫ, УСТОЙЧИВЫХ К ДЕЙСТВИЮ УФ-Б РАДИАЦИИ Воздействие достигающей поверхности Земли ультрафиолетовой области солнечной радиации (УФ-радиации) на растительные объекты привлекает пристальное внимание ученых в связи с изменением состояния озонового слоя атмосферы [4, 8]. Значительное его утоньшение отмечено для многих континентов планеты [4], что является следствием техногенных процессов [17]. Это, в свою очередь, приводит к тому, что на поверхности Земли возрастает уровень излучения в диапазоне длин волн от 280 до 320 нм, который относится к ультрафиолетовой области спектра , а именно - к его "среднему" или "Б"-участку, обозначаемому как как "УФ-Б радиация" [4, 8]. Именно озон поглощает практически всю УФ-радиацию ниже 280 нм [6].
УФ-радиация является одним из сильных стрессовых факторов окружающей среды, и понимание того, каким образом живые организмы реагируют на него, позволит оценить опасность ее воздействия. Показано, что УФ-радиация вызывает морфо-физиологические изменения у растений. Она снижала линейный рост стебля, уменьшала коэффициент кущения и число колосоносных побегов у пшеницы [19]. У проростков картофеля увеличение продолжительности ежедневного действия УФ-Б радиации приводило к уменьшению высоты стебля и поверхности листьев [5]. При этом эффекты действия УФ-Б радиации зависели от стадии роста растений. Наибольшая чувствительность к действию радиации отмечалась при переходе от вегетативной и репродуктивной стадии развития [9, 10, 14]. Отмечено также изменение гормонального статуса растений при действии УФ-Б радиации. Так, например, у растений арабидопсиса отмечалось увеличение накопления абсцизовой кислоты [15]. Все это приводит к образованию растений с заторможенным ростом, плохой пигментацией и нарушениям формирования листового аппарата. Это, в свою очередь, отражалось и на функционировании их фотосинтетического аппарата. При этом фотосистема II более чувствительна к действию УФ-Б радиации, чем фотосистемы 1. Это обусловлено тем, что УФ-облучение подавляет фотохимическую активность фотосистемы II, вызывая уменьшение потока электронов между фотосистемами [18].
Ультрафиолетовые лучи индуцируют также различные повреждения ДНК, при этом пиримидины в 10 раз более чувствительны к действию ультрафиолета, чем пурины [11]. Ультрафиолетовое излучение, действуя на ДНК, может также индуцировать большое число хромосомных аберраций в клетках растений [3]. Все это может быть причиной повышения уровня мутаций в растительных клетках, что, в свою очередь, негативно сказывается на сохранении генофонда живых организмов.
В связи этим изучение действия УФ-Б радиации на растительные объекты и получение устойчивых к этому фактору культур имеет важное народно-хозяйственное значение.
Основной зерновой культурой сельского хозяйства России является пшеница, с которой давно ведется селекционная работа с использованием традиционных подходов. В последнее время все более широкое применение находят также и биотехнологические приемы на основе селекции растительных клеток в условиях in vitro [12]. Такой подход существенно ускоряет селекционный процесс в силу реализации свойственной растительной клетке тотипонентности.
В связи с этим, целью настоящего исследования являлось получение каллусных культур пшеницы, обладающих повышенной устойчивостью к действию ультрафиолетового облучения и изучение их морфо-физиологических характеристик.
МАТЕРИАЛЫ И МЕТОДЫ
В качестве объекта исследования использовали каллусные культуры, инициированные из двух генотипов яровой мягкой пшеницы Triticum aestivum - сорта Таежная и линии Фотос.
Сорт Таежная был выведен в Красноярском НИИСХ методом индивидуального отбора из третьего поколения гибридной комбинации К-441 (Л-330 х М-341). Разновидность лютесценс. Достоинством сорта является скороспелость (созревает раньше стандарта на 2-11 дней), устойчивость к полеганию (оценивается в 4,9 балла, выше стандарта на 1,3 балла) и к прорастанию на корню и в валках. Дает семена с высокой послеуборочной всхожестью.
Линия Фотос (Л 40959) была получена индивидуальным отбором из гибридной популяции от скрещивания сортов Московская 35 и Жница в Башкирском НИИЗиС. Разновидность мильтурум. По качеству зерна относится к ценным пшеницам. Линия устойчива к полеганию, среднеранняя - созревает одновременно с сортом Жница (вегетационный период - 85-90 дней). Проявляет высокую устойчивость к поражению мучнистой росой и бурой ржавчиной, пыльной головней поражается слабо. Обладает хорошей комбинационной способностью.
Для инициации каллусной ткани семена пшеницы стерилизовали 0,1% сулемой в течение 10 минут, заливали небольшим количеством стерильной воды и через 16-24 часа вычленяли из них зародыши. Материал помещали в чашки Петри диаметром 9 см по 12-18 штук, щитком вниз, на поверхность модифицированной агаризованной питательной среды Мурасиге-Скуга [7], содержащей 2,5 мг/л 2,4-дихлорфеноксиуксусной кислоты, 20 мг/л аскорбиновой кислоты и 10 мг/л AgNO3 и культивировали при температуре 20-22oС и 16-час. фотопериоде (интенсивность освещенности 4000 люкс). Длительность пассажа составляла 28 дней.
В качестве источника УФ-Б радиации использовали лампу ДРЛФ-400 с удаленной внешней колбой, являющейся эквивалентом бактерицидной ртутной лампы высокого давления типа ПРК-2 (ДРТ). Для селекции клеточных линий пшеницы устойчивых к действию УФ-Б радиации растительный материал культивировали в стеклянных чашках фирмы "Anumbra" (Германия), не пропускающих излучение ниже 280 нм.
Интенсивность облучения (1,66; 1,11; 0,74; 0,55; 0,46 и 0,39 Вт/м2 по УФ-Б) регулировали изменением расстояние чашек с культурами от лампы. Время облучения составляло по 2 часа каждый день (с 16 до 18 час.). Контрольные варианты были изолированы от лампы обычным стеклом.
Исследовалось действие УФ-Б на процесс каллусообразования, интенсивность роста и регенерационную способность неорганизованно растущей каллусной ткани яровой мягкой пшеницы T. aestivum сорта Таежная и линии Фотос, а также на морфологические характеристики клеточных линий.
Анализировали прирост каллусной массы путем индивидуального взвешивания каллусных агрегатов в стерильных условиях на торсионных весах в момент очередного пассирования. Результаты оценивали в конце каждого пассажа по количеству образовавшихся каллусов из первичного экспланта и по их массе, которая учитывалась индивидуально для каждого экспланта, что позволило выделить клеточные линии, ведущие свое происхождение от одного первичного экспланта.
Для изучения морфологии клеток каллусные ткани замораживали, используя для этого микротом-криостат, готовили тонкие срезы, которые сразу же окрашивали малахитовым зеленым [16]. Анализировали размеры клеток и толщину клеточных стенок на микроскопе "Ergavall" (Германия) с помощью окуляр-микрометра.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Для осуществления поставленной задачи необходимо получить растущие в условиях in vitro каллусные культуры пшеницы обладающие устойчивостью к действию УФ-Б радиации. Для этого каллусные ткани выращивали в условиях УФ-облучения в течение ряда субкультивирований, а затем был произведен отбор устойчивых клеточных линий.
Каллусные культуры пшеницы (сорт Таежная и линия Фотос) представляли собой агрегаты светло-желтого цвета, никогда не зеленеющие, плотные, часто с выраженной внутренней структурой, обычно нескольких, хорошо различимых типов, соотношение которых зависело от условий культивирования, возраста культуры, частоты пассирования и индивидуальной массы каллусных агрегатов.
УФ-Б радиация (1,66 и 0,74 Вт/м2 по УФ-Б) негативно влияла как на интенсивность формирования каллусов на эксплантах, так и на их массу (табл.1, рис. 1). При этом у линии Фотос интенсивность каллусообразования в условиях действия УФ-Б радиации была почти в три раза ниже, чем в контроле (табл.1). У сорта Таежная эти изменения были выражены в значительно меньшей степени. Следовательно, изученные генотипы отличались по ответной реакции на действие УФ-Б радиации.
Ингибирующее действие радиации прослеживается и при определении массы сформировавшихся каллусов. При этом у обоих генотипов увеличение дозы УФ-Б сопровождалось линейным снижением этого показателя (табл.1). Так при интенсивности 1,66 Вт/м2 по УФ-Б масса каллуса линии Фотос составляла в среднем 51,2% от контроля, а Таежной - 45,0%. А при интенсивности 0,74 Вт/м2 - 65,3% и 64,2% соответственно.
Таким образом два изученных генотипа пшеницы отличались по реакции на действие УФ-Б радиации. У линии Фотос облучение в большей степени влияло на инициацию каллусов и в значительно меньшей степени на последующий их рост. У сорта Таежная процент каллусообразования в присутствии УФ-Б оставался довольно высоким, тогда как масса образовавшихся калусов была ниже, чем у линии Фотос (при одинаковой дозе радиации).
Аналогичные закономерности отмечались и при более длительном действии УФ-Б радиации на каллусные культуры (рис. 2). Так, каллусная ткань выращиваемая в контрольных условиях или в условиях облучения интенсивностью 0,74 Вт/м2 по УФ-Б сохраняла жизнеспособность и имела индекс роста порядка 2,1-2,6. В то время как каллусы, выращиваемые при более высокой интенсивности облучения (1,66 Вт/м2 по УФ-Б) прекращали рост в течение первого месяца культивирования. При это культуры инициированные из сорта Таежная имели несколько больший прирост каллусной массы и индекс роста, чем культуры инициированные из линии Фотос.
При дальнейшем культивировании выяснилось, что выбранные дозы оказались летальными для обоих генотипов, поскольку наблюдалось только образование первичного каллуса из тканей экспланта. При их дальнейшем культивировании в контрольных условиях (без облучения УФ-Б) прироста каллусной массы не наблюдалось, хотя по внешнему виду культуры сохраняли характерный для живой ткани светло-желтый цвет и консистенцию. Кроме того УФ-Б радиация полностью блокировало морфогенетические процессы в культурах. В результате этого в последующих опытах условия селекции были несколько изменены - уменьшена общая доза радиации и введена более дифференцированная шкала (1,11; 0,74; 0,55; 0,46 и 0,39 Вт/м2 по УФ-Б). Было выяснено, что доза 1,11 и 0,74 Вт/м2 УФ-Б оказывала летальное действие на каллус сорта Таежная: после месяца культивирования при облучении сохранили рост 22,7% экспланта, после второго месяца - 0% (рис.2). При дозе 0,55 Вт/м2 после 2 субкультивирований были выделены 11,2% жизнеспособных колоний, которые давали высокий прирост массы. Линия Фотос оказалась менее подверженной действию УФ-Б. Даже при дозе 1,11 Вт/м2 по УФ-Б наблюдался некоторый прирост массы - 27,6% после 1 месяца и 18,9% после второго (рис.2).
В результате проделанной работы для линии Фотос удалось выделить несколько клеточных линий, способных сохранять рост на уровне контроля в присутствии УФ-Б радиации. Изучение морфологических характеристик показало, что все культуры состоят из клеток паренхимного типа имеющих крупную вакуоль, занимающую большую ее часть (рис.3). Размеры клеток и их структура у полученных в результате селекции линий и контрольных вариантов каллусных культур несколько отличались. Так, у культур подвергнутых действию УФ-Б радиации размеры клеток были несколько меньше (табл.2), а толщина клеточных стенок больше (табл.3) по сравнению с контролем. Следует также отметить, что чем выше доза действия УФ-Б радиации на культуры, тем толще их клеточные стенки.
Все это свидетельствует о том, что УФ-Б радиация вызывает изменения в морфо-физиологических характеристиках клеток пшеницы, выращиваемых в условиях in vitro. В первую очередь, это подавление роста - растяжением и, как следствие, утолщение клеточных стенок, а, во-вторых, ослабление уровня внутритканевой дифференциации. Из этого мы можем заключить, что реакция клеток пшеницы, выращиваемых в условиях in vitro, на действие УФ-Б радиации в определенной степени аналогична таковой интактных растений [19], что свидетельствует о перспективности использования таких биотехнологических подходов с целью ускорения селекционного процесса в отношении важнейших для России сельскохозяйственных культур, в том числе и пшеницы.
ВЫВОДЫ
П.В.Лапшин, Н.В.Трошенкова, Г.А.Дубравина*, Н.В.Загоскина*, Р.Г.Бутенко*
Московская сельскохозяйственная академия им. К.А.Тимирязева, кафедра сельскохозяйственной биотехнологии;
*Институт фзиологии растений им. К.А.Тимирязева РАН, Москва, 123276, Ботаническая 35

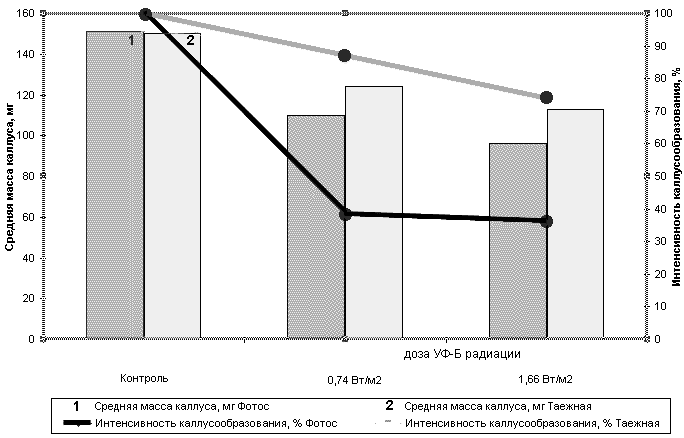
Рис.1. Интенсивность каллусообразования и прирост массы у каллусов яровой мягкой пшеницы сорта Таежная и линии Фотос при действии различных доз УФ-Б радиации.
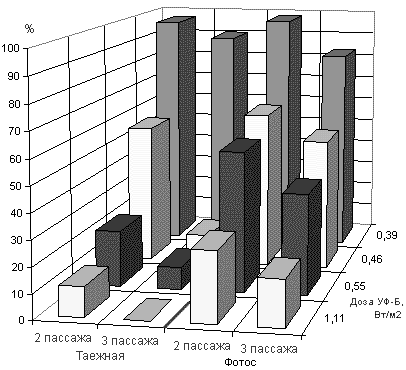
Рис. 2. Влияние длительного воздействия различных доз УФ-Б радиации на рост каллусных культур яровой мягкой пшеницы сорта Таежная и линии Фотос.

Рис 3. Каллусная культура пшеницы линии 60/5. Окрашивание малахитовым зеленым. Возраст ткани 20 дней.


Помимо паренхимных клеток в каллусных культурах довольно часто наблюдалось формирование меристематических очагов. Более интенсивно этот процесс происходил в контрольных вариантах, а также в культурах, полученных в результате селекции в условиях действия низких доз УФ-Б радиации. Меристематические клетки обычно имели округло-овальную форму, густую цитоплазму, крупное ядро и тонкую клеточную стенку (рис.4). Во многих линиях встречались также и трахеидальные элементы (как на начальной стадии формирования, так и в виде сосудистых пучков) (рис.5).

Рис 4. Меристематические очаги в каллусной культуре пшеницы (окрашивание флорглюцином).

Рис 5. Трахеидальные элементы в каллусной культуре пшеницы (окрашивание малахитовым зеленым).
ЛИТЕРАТУРА
|
|






