Происхождение, развитие и размножение химер
проф. Р. Лайнбергер
факультет растениеводства, Texas A&M University
ВВЕДЕНИЕ
Растение считается химерой, когда в растущих тканях совместно присутствуют клетки более чем одного генотипа. Вариегатные растения - возможно наиболее частый тип химер и, конечно, это наиболее удобный пример для представления основной концепции. Клетки в вариегатном листе происходят из апикальной меристемы побега, но некоторые клетки отличаются неспособностью синтезировать хлорофилл. Они проявляются как более белые, чем остальные даже при том, что являются частью той же самой ткани. Много важных сортов декоративнолистных и красивоцветущих садовых растений - химеры. Дерен белый (Cornus alba) сорт 'Argenteo Marginata', барвинок малый (Vinca minor) 'Variegata', живучка ползучая (Ajuga reptans) 'Burgundy Glow', и много сортов хост (Hosta), диффенбахий (Diffenbachia), пеперомий (Peperomia), хлорофитумов (Chlorophytum), и африканских фиалок (Saintpaulia) - это малая толика химер, которые имеют вариегатную листву.
Цель этой статьи - исследовать происхождение и развитие химерных растений, назвать узкие места, которые сопровождают размножение химер, и обсуждать существенные для цветоводства химеры, другие чем вариегатные по типу листвы.
КОНЦЕПЦИЯ АПИКАЛЬНОЙ ОРГАНИЗАЦИИ
Обсуждение происхождения химер не было бы закончено без обзора организации вершины (апекса) побега. Пример клеточного деления, частота деления, и организация слоев клеток в апексе связаны в определении получаемого типа химеры и ее стабильности в результате.
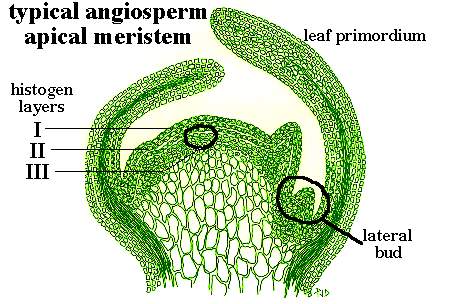
Апикальная меристема побега - место, где находятся большинство клеток, которые производят все тело растения. Клеточные деления происходят в активно растущем побеге очень быстро, и поделившиеся клетки растут в длину и ширину, обечпечивая рост побега в длину. Древесные и некоторые травянистые растения содержат вторичные меристемы, которые ответственны за увеличение толщины побега, но этот вторичный рост не будет рассматриваться здесь. На рис. 1 - схема поперечного разреза апикальной меристемы типичного древесного или травистого двудольного растения. Примордиальные листья возникают по сторонам от апикального купола, и боковые почки развиваются в аксаллах этих молодых листьев.
Апекс состоит из внешней слоистой области (туника) и внутренней области, где иерархия клеток не очевидна (корпус). Упорядоченная организация клеточных делений в тунике приводит к поддержке дискретных слоев, с количеством слоев, изменяющимся в зависимости от вида. Важно заметить, что слои сохраняют свою организацию на области, из которых формируются листья и боковые почки.
Клетки самого внешнего слоя (L.I), формируют эпидермис (см. рис. 1). Эпидермальный слой непрерывен как внешняя оболочка у всех тканей листа, стебля, цветочных лепестков и т.д. Производные второго слоя (L.II) дают несколько слоев в пределах стебля и большую пропорцию клеток в листовой пластинке. Производные третьего слоя (L.III) ответственны за большинство внутренних тканей стебля и сосудов в листьях. Значение клеточных слоев в формировании органов будет обсуждено более подробно ниже.
ПРОИСХОЖДЕНИЕ ХИМЕР
Химеры возникают когда клетка подвергается мутации. Эта мутация может быть самопроизвольной, или может быть вызвана радиацией или обработкой с химическими мутагенами. Если клетка, которая изменяется, находится около вершины апикального купола, то все другие клетки, которые происходят от нее, будут также иметь видоизмененный тип. В результате клетки различных генотипов будут расти вместе в ткани растения. Это и есть определение химеры.
Если локализация клетки в меристеме в момент мутации такова, что она будет делиться небольшое количество раз, то вероятность визуального обнаружения этой мутации в целом растении низка. Кроме того, если мутация дает фенотип, который не очень отличается морфологически от остального растения, тогда вероятность идентификации химерности у такого растения также небольшая. Мутация, которая дает обесцвечивание хлорофилла (вариегатность), легко обнаруживается.
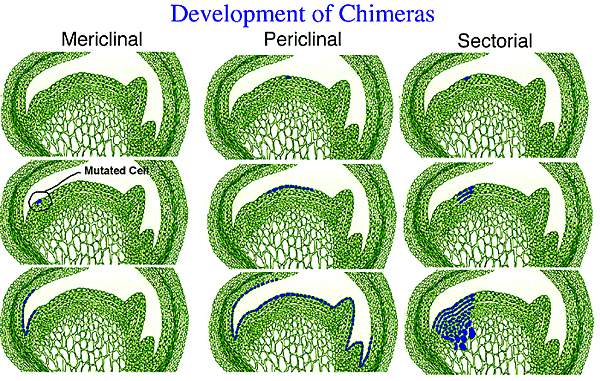
Периклинальные, мериклинальные и секторные химеры
Химерные растения могут быть систематизированы на основе размещения и пропорции измененных клеток к неизмененным в апикальной меристеме. Периклинальные химеры (см. рас. 2) - наиболее важный тип, так как они относительно устойчивы и могут быть вегетативно размножены. Мутация дает периклинальную химеру, если мутировавшая клетка находилась около апикального купола и так, что образующиеся от нее клетки сформировали полный слой из измененного типа. В результате меристема содержит один слой, который генетически отличается от остальной меристемы. Если, например, мутация происходит в L.I, то эпидермальный слой побега, который вырастет после мутации будет нового генетического типа.
Классический пример периклинальной химеры L.I - ежевика без шипов. Эпидермальный слой у этого типа не производит никаких шипов. Эпидермис без шипов покрывает остальные слои стебля, чей генотип содержит информацию для образования колючек. Что может быть продемонстрировано индукцией побегов от корней. Адвентивные побеги, которые образуются на корневых черенках, нехимерны и поэтому возвращаются к исходному генотипу с шипами.
Мериклинальные химеры (рис. 2) получаются, когда потомки измененной клетки неполностью накрывают апикальный купол. Измененный клеточный слой может поддерживаться только на одной стороне меристемы. В результате химерные побеги или листья развиваются только с одной стороны оси побега, в то время как те, которые дифференцируются на другой стороне меристемы будут нормальными, нехимерными. Большинство клеток в меристеме мериклинального типа формируют сравнительно небольшую долю клеток, которые можно обнажужить только на маленькой части одного листа. Также как и в случае с периклинальными химерами, мериклинальные химеры ограничиваются одним клеточным слоем.
Секторные химеры (рис. 2) получаются из мутаций, которые затрагивают целый сектор апикальной меристемы, простирающийся через все клеточные слои вглубь. Этот тип химер непостоянен и может давать нехимерные побеги и листья. И нормальный и химерный типы могут быть получены, в зависимости от места на апексе от которого дифференцируется конкретный лист или побег.
РАСПОЗНАВАНИЕ ХИМЕРНЫХ ЭКЗЕМПЛЯРОВ
Осторожное наблюдение экземпляра с вариегатной окраской листьев у двудольных позволяет частично понять природу химерных слоев в апексе побега. Полная характеристика всех трех слоев не может быть сделана визуально, так как большинство клеток эпидермального слоя листа у двудольных не производит хлоропласты, даже если генетическая информация для развития хлоропласта в них присутствует. Однако, если меристема содержит хлоропласты в L.I, но они должны наблюдаться при микроскопическом анализе.
С точки зрения терминологии, природа слоев клеток апекса, описывается с использованием латинских символов в верхнем регистре и нужно сделать попытку обозначить генетическую композицию клеток, которые там содержатся. Например, правильное обозначение для периклинальной химеры с генетически зеленым L.I, белым L.II, и зеленым L.III - G-W-G.
В листе двудольных, край листа формируется из L.II. Доля края листа, сформированная за счет L.II варьирует от вида к виду, и до некоторой степени меняется и среди листьев одного растения. Лист, изображенный на рисунке 2A был произведен меристемой, которая была G-G-G (нехимерная, не вариегатная), в то время как лист на рисунке 2B был произведен G-W-G вариегатным спортом.
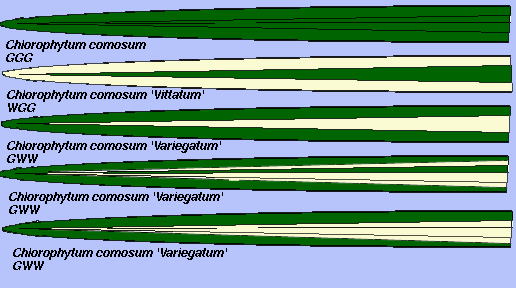
У видов однодольных слой L.I вносит вклад в развитие края, а большая часть центральной части листа развивается из L.II. В отличие от двудольных, L.III вносит только маленькую долю клеток в листовую пластинку. На рис. 3 A-E представлены типы вариегатности у листьев хлорофитума (Chlorophytum comosum). Рис. 3A, сверху - G-G-G (нехимерный, не вариегатный), рис. 3B, второй сверху - тип W-G-G, и на рис. 3C (в середине) - тип G-W-W. Вариабильность листьев и вклад L.I и L.II в соотношение клеток в листьях индивидуального растения показано на рис. 3D и 3E (два нижних). Оба листа были взяты от одного растения, но зененая окантовка от L.I у листа на рис. 3D была намного больше. Такие наблюдения показывают, варьирование вклада различных слоев меристемы в развитие разных листьев и что это жестко не детерминировано.
ПРОЯВЛЕНИЕ ДРУГИХ ХИМЕР КРОМЕ ВАРИЕГАТНОСТИ
В то время как вариегатность представляет хорошо видимое проявление химерности, из предыдущего обсуждения должно быть очевидно, что теоретически растение может быть химерным фактически по любому признаку. Другим частым примером визуально хорошо заметной химерности является потеря эпидермальных выступающих частей (ежевика без шипов, голые ("fuzzless") персики), сдвиг в цвете у прицветниковых листьев Poinsettia, и вариации цветовой палитры у гвоздик и хризантем.
Химеры по уровню плоидности интенсивно изучились на плодовых культурах. Тетраплоиды (удвоение нормального числа хромосом) дает плоды, которые могут быть намного больше чем нормальные, когда эта черта зафиксирована в L.II и L.III. Это явление изучилось на яблонях и винограде.
РАЗМНОЖЕНИЕ ХИМЕРНЫХ РАСТЕНИЙ
Мериклинальные и секторные химеры - по своей природе очень нестабильны и вероятность размножения растений с тем же самым фенотипом от этих типов низка. Периклинальные химеры очень устойчивы и в некоторых случаях (как у вариегатного Chlorophytum) химерное растение - наиболее доступная форма в торговле.
Наиболее хорошо при ветативном размножении сохраняются свойства периклинальных химер. Эта свойство идет от того, что расположение клеточных слоев в апикальной меристеме боковых побегов является идентичным расположению тих слоев в меристеме исходного побега. Таким образом слои I, II и III вносят свои клетки в дифференцирующиеся боковые меристемы. Укоренененные черенки, глазков, прививка и деление куста - все эти методы размножения дают высококачественное воспроизведение. Важным исключением из этого правила является дифференциация побегов от адвентивных почек. Случай, который это иллюстрирует, был описан выше - ежевика без шипов. Адвентивные корни, которые формируются на стеблевом черенке ежевики, происходят из субэпидермальных тканей стебля (L.II и L.III). Если получить корневую поросль от этих растений, адвентивные побеги не будут содержать мутантный слой (L.I) "без шипов" и побеги будут нормально околючены.
Африканские фиалки с химерным полосатым цветком (тип "pinwheel") (Фото 4) аналогично не могут быть размножены с сохранением этого типа окраски цветка через листовые черенки. Побеги, которые происходят от листовых черенков будут иметь или какой-то один цвет или различные спорты с нерегулярными пятнами двух цветов: Фото 5), не соответстующие сорту. Размножение химерных сортов африканских фиалок с полосатыми цветками в настоящее время достигается отдением боковых побегов из боковых почек. Это иллюстрирует, что палитра цветка определяется распределением клеточных слоев в апикальной меристеме.
РАЗДЕЛЕНИЕ ХИМЕР В КУЛЬТУРЕ ТКАНЕЙ
Культура тканей растений позволяет размножать растения от очень маленьких фрагментов ткани и в некоторых слуяаях даже от единичных изолированных клеток (культура протопластов). Когда эти методы применяются на химерных растениях, технология культуры тканей предлагает беспрецедентный инструмент для "разбиения" или разделения химер для морфологического изучения составляющих их генотипов. Факт, что химеры могут разелиться при формировании адвентивных почек с сопутствующим нарушением расположения слоев в верхушечной меристеме, создает серьезные проблемы в использовании этой технологии для быстрого клонального размножения.
Работы в нашей лаборатории показали, что живучка ползучая (Ajuga reptans) сорта 'Burgundy Glow' размножается in vitro и от пазушных почек и от адвентивных побегов: Фото 6. Те растения, которые происходят из пазушных почек, соответствуют сорту 'Burgundy Glow'. Однако, вырастают также и нетипичные всходы. Они имеют два типа. Наиболее частый тип - "bronze" с равномерно глубоким зеленым типом листвы. Другой тип полностью лишен хлорофилла, он розовый в культуре, но при попытке укоренения он гибнет из-за неспособности к фотосинтезу. Основное размножение происходит через развитие пазушных почек с почти 70% воспроизведением сорта 'Burgundy Glow': Фото 7.
РЕЗЮМЕ
Устойчивые периклинальные химеры у растений это спорты с уникальными декоративными характеристиками. Способность размножать эти спорты с сохранением их свойств зависит от использования методики размножения: через боковые почки. Слоистая организация апикальной меристемы, которая является характерной для периклинальной химеры, поддерживается и в боковых меристемах, но обычно теряется при дифференцировке адвентивных побегов. Методология культуры тканей растения позволяет разделять химерные генотипы у некоторых видов, облегчая изучение составляющих генотипов. Эта особенность культуры тканей может быть ограничением в системах, размножающихся in vitro через формирование побегов из адвентивных почек. В этом случае может происходить получение нетипичных растений.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
- Dermen, H. 1955. A 2-4-2 chimera of McIntosh apple. J. Wash. Acad. of Sci. 45:324-327.
- Dermen, H. 1960. Nature of Plant Sports. The Horticultural Magazine, July 1960, pp. 123-173.
- Esau, K. 1965. Plant Anatomy. Second Edition, John Wiley & Sons, Inc., New York, pp. 89-112.
- Stewart, R.N. and H. Dermen. 1979. Ontogeny in monocotyledons as revealed by studies of the developmental anatomy of periclinal chloroplast chimeras. Amer. J. Bot. 66, pp. 47-58.