СОДЕРЖАНИЕ ФОТОСИНТЕТИЧЕСКИХ ПИГМЕНТОВ У ЦВЕТНЫХ ФОРМ КАКТУСОВ В СРАВНЕНИИ С НЕКОТОРЫМИ ДРУГИМИ СУККУЛЕНТНЫМИ РАСТЕНИЯМИ
И.М.Скулкин, кандидат биологических наук, доцент; Е.П.Артемьева
фото 1.
В 1935-1941 гг. японским кактусоводом Ватанабе впервые была получена первая устойчивая аномальная цветная форма (АЦФ) у кактуса Gymnocalycium mihanovichii var. friedrichii. В последующие годы в разных странах были получены сотни АЦФ разных видов кактусов, окрашенных в различные оттенки красного и желтого цвета, а также белые, коричневые, трехцветные и пр. Все эти формы справедливо рассматриваются как хлорофильные мутанты, то есть растения с глубокими нарушениями различных этапов синтеза основного фотосинтетического пигмента - хлорофилла, поэтому их выращивают почти исключительно в привитом виде, чаще всего на подвое Hylocereus triangularis.
фото 2.
Причины возникновения хлорофильных мутантов весьма разнообразны. Они могут возникать в результате: 1) мутаций генов и изменения структуры хромосом (ядерные мутации); 2) мутаций в ДНК хлоропласта (пластидные мутации); 3) изменения ДНК цитоплазмы (цитоплазменные мутации). Среди этих различных генетических типов - ядерных, пластидных (пластомных), цитоплазменных - имеются фенотипически сходные, то есть неотличимые по внешним признакам мутанты [7].
У хлорофильных мутантов не просто уменьшено количество зеленых и желтых пигментов, но при этом изменяется и их соотношение.
фото 3.
Кактусоводы-любителий России неоднократно отмечали, что АЦФ появляются в значительно большем количестве при продолжительном пребывании в условиях очень яркого света и экстремально высоких температур. Однако, конкретный механизм "запуска" мутантных генов у кактусов в экстремальных условиях пока не изучен.
С сожалением констатируем, что сих пор в отношении названий АЦФ кактусов до сих пор не выработана единая номенклатура, поэтому в разных странах одни и те же формы, преимущественно японской селекции, фигурируют либо под оригинальными японскими же названиями типа "Hibotan", "Nishiki", либо под "европеизированными" названиями "Ruby Ball", "Black Botan", а чаще всего распространяются вообще без всяких названий. Весьма показательно то, что даже в Голландии, где давно налажено массовое производство АЦФ, в каталогах цветоводческих фирм они чаще всего обозначаются лишь ботаническим названием исходного рода - "Gymnocalycium", то есть Gymnocalycium mihanovichii var.friedrichii, без названия конкретной АЦФ.
фото 4.
Основной целью работы являлась проверка рабочей гипотезы, являются ли все АЦФ кактусов истинными "бесхлорофильными" мутантами. Задачами исследования были: 1) определение содержания фотосинтетических пигментов у АЦФ; 2) сравнение количества хлорофиллов и каротиноидов у АЦФ, подвоев, листовых суккулентов и обычных растений; 3) оценка соотношений "хлорофиллы : каротиноиды" и "хлорофилл a : хлорофилл b" у АЦФ в сравнении с тремя контрольными группами.
МАТЕРИАЛ И МЕТОДИКА. Летом 2000 г. в Ботаническом саду УрГУ изучалось содержание фотосинтетических пигментов - хлорофиллов a и b, а также каротиноидов - у 9 различных АЦФ кактусов, в том числе Eriocactus magnificus f. aurea, Chamaecereus silvestrii f. aurea, у шести форм Gymnocalycium mihanovichii var. friedrichii. Поскольку единой общепринятой системы названий АЦФ не существует, мы сочли возможным использовать собственные условные названия, характеризующие окраску АЦФ на момент исследования, как-то:
PURPLE - насыщенно малиновая форма (фото1);
RED - яркая ало-красная форма (фото 2);
PINK - яркая розовая форма (иногда называемая "Pottii") фото 3;
ROSE - бледно-розовая форма с редкими темными пятнами (фото 4);
MULTICOLOR - трехцветная желто-красно-зеленая пятнистая форма (вероятнее всего, это "Hibotan") фото 5;
BLACK - темно-коричневая форма с розоватыми вкраплениями (вероятнее всего, "Black Botan") фото 6.
фото 5.
В эту же группу был включен и корнесобственный Gymnocalycium mihanovichii var. friedrichii s.str. - вид, на основе которого было получено большинство АЦФ гимнокалициумов.
Для сравнения были взяты 7 видов кактусов, обычно используемых в качестве подвоев (Hylocereus, Selenicereus и др.), 18 видов суккулентов из семейства Crassulaceae, а также 3 вида несуккулентных растений (ландыш, кислица, амарант). Все растения, кроме последней группы, в течение двух месяцев перед исследованиями находились в теплице при температурах около +30 °С в условиях хорошего естественного освещения.
фото 6.
Для исследования брались исключительно наружные участки коры кактуса вместе с эпидермисом толщиной 1,5-2 мм, поскольку основная часть пигментов у кактусов сосредоточена именно в этой зоне. У листовых суккулентов брали преимущественно участки из средней части целого листа. Образцы фиксировались, измельчались, затем пигменты экстрагировались 85 %-ным ацетоном. Концентрацию пигментов определяли на спектрофотометре "Ломо" СФ-46. Повторность определения для всех видов была трехкратной.
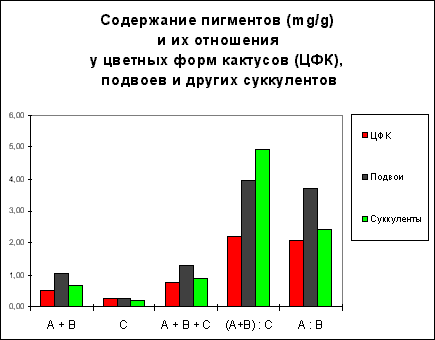
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ. Как и ожидалось, из четырех групп изученных нами растений наименьшее количество хлорофиллов оказалось у АЦФ кактусов (табл.1). Однако удивительно, что у обычных суккулентов из семейства толстянковых хлорофиллов оказалось ненамного больше. У подвоев количество зеленых пигментов больше примерно в два раза, а у обычных лиственных растений, в основном тенелюбивых, содержание хлорофиллов выше на порядок (12,89 против 0,47-1,04 мг/г сырого веса, соответственно), что вполне согласуется с литературными данными.
Таблица 1
ГРУППЫ
РАСТЕНИЙ
A
+ B
C
A
+ B + C
(A+B)
: C
A
: B
АЦФ
кактусов
0,47
0,25
0,73
2,07
1,98
Кактусы,
используемые для подвоев
1,04
0,26
1,30
3,96
3,72
Суккулентные
растения семейства толстянковых
0,67
0,21
0,88
4,94
2,42
Несуккулентные
растения (контрольная группа)
12,89
2,82
15,71
4,52
4,03
Содержание пигментов (мг/г сырого веса) и их соотношения у суккулентных и несуккулентных растений
Обозначения: хлорофилл a + хлорофилл b (А+В) ; каротиноиды (С); хлорофиллы + каротиноиды (А+В+С); отношение "хлорофиллы : каротиноиды" (А+В):С; отношение "хлорофилл a : хлорофилл b" (А : В)
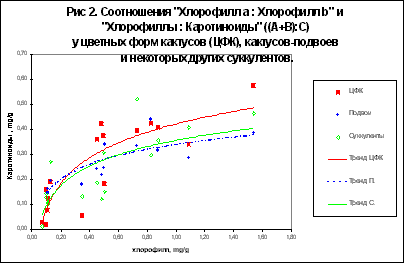
Наибольший интерес представляло сравнение содержания желтых и красных пигментов, то есть каротиноидов, у цветных кактусов с другими группами растений, однако различий между АЦФ, подвоями и толстянковыми практически не выявлено. Из этого однозначно следует, что окраска цветных форм кактусов не связана с повышенным содержанием каротиноидов по сравнению с другими суккулентами. У обычных растений содержание каротиноидов на порядок выше (2,82 мг/г), отсюда и сумма пигментов (А+В+С) также выше в 10-20 раз по сравнению с цветными формами и другими суккулентами.
фото 7.
Отношение "хлорофиллы : каротиноиды" оказалось наименьшим у цветных форм кактусов, что выглядит естественным, поскольку хлорофиллов у них меньше, а каротиноидов примерно столько же, как и у других суккулентов. Различия между подвоями, толстянковыми и лиственными растениями по этому показателю, были незначительными, варьируя от 3,96 до 4,94.
У растений открытых, сильно освещенных местообитаний (гелиофитов), отношение "хлорофиллы : каротиноиды" обычно больше, чем у теневыносливых растений затененных местообитаний (сциофитов), в среднем 5,3-5,5 против 3,6-4,6. При сопоставлении наших результатов с этими данными становится ясно, что АЦФ кактусов имеют вообще исключительно низкие значения этого показателя, подвои приближаются скорее к теневыносливым растениям, а толстянковые при всей своей очевидной приспособленности к экстремально сильному освещению занимают, как ни странно, лишь промежуточное положение между гелиофитами и сциофитами!
Отношение "хлорофилл а : хлорофилл в", у АЦФ по сравнению с кактусами-подвоями и лиственными растениями было существенно меньшим, хотя и незначительно отличалось по сравнению с толстянковыми (2,07 против 2,42).
У гелиофитов форма "a" значительно преобладает над формой "b", а в затенении доля хлорофилла "а" падает и отношение а:b гораздо меньше. Cоотношение пигментов меняется в течение онтогенеза пластиды, листа и всего организма, но оно относительнo определенно и генетически детерминировано. Отмечается также, что отношение "a : b" составляет в среднем 3,0 [6]. У цветковых альпийских растений, типичных гелиофитов, отношение "a : b" в среднем составляет 5,6, в то время как растения "зеленой" тени, то есть сциофиты в нижних ярусах растительных сообществ, имеют этот показатель на уровне 2,6, а погруженные водные растения - 2,3.
фото 8.
Сравнение этих данных с нашими результатами показывает разительное несоответствие: 1) АЦФ кактусов и растения семейства толстянковых гораздо ближе к сциофитам, нежели к гелиофитам; 2) кактусы-подвои так же ближе к сциофитам; 3) взятые для контроля 3 вида лиственных растений, из которых два являются заведомыми сциофитами (ландыш и кисличка) дают среднее значение по этому показателю 4,03, то есть существенно выше, чем 3,72 у подвоев и 2,42 у типичных гелиофитов- толстянковых. На данный момент мы затрудняемся объективно интерпретировать эти результаты.
При сравнительном анализе изученных показателей у различных АЦФ открывается еще более удивительная картина (табл.2).
Таблица 2
Содержание пигментов и их соотношения у цветных форм кактусов и исходного вида Gymnocalycium mihanovichii var. friedrichii s.str. (ранжированы по содержанию хлорофиллов А+В)
№
Форма
Часть
растения
А+В
С
А+В+С
(А+В):С
А:В
1
MULTICOLOR
(HIBOTAN)
черная
детка 7 мм
1,54
0,58
2,12
2,55
2,15
2
MULTICOLOR
(HIBOTAN)
бутон
1,09
0,34
1,43
3,20
1,97
3
BLACK
молодые
ткани
0,88
0,41
1,29
2,15
3,04
4
MULTICOLOR
(HIBOTAN)
оранжевая
детка 7 мм
0,83
0,43
1,25
1,94
0,67
5
MULTICOLOR
(HIBOTAN)
молодые
ткани
0,73
0,39
1,13
1,63
2,32
6
BLACK
бутон
0,50
0,18
0,69
2,73
1,88
7
MULTICOLOR
(HIBOTAN)
детка
15 мм
0,50
0,38
0,87
1,27
4,24
8
MULTICOLOR
(HIBOTAN)
детка
15 мм
0,48
0,42
0,91
1,05
3,70
9
Gymnocalycium
friedrichii s.str.
сеянец
1 год
0,45
0,36
0,82
1,25
0,53
10
Eriocactus
magnificus AUREA
молодые
ткани
0,35
0,06
0,41
6,08
3,05
11
PURPLE
детка
7 мм
0,13
0,19
0,33
0,66
0,53
12
Chamaecereus
silvestrii AUREA
молодые
ткани
0,11
0,12
0,23
0,86
3,15
13
ROSE
детка
15 мм
0,11
0,08
0,19
1,12
1,11
14
PINK
детка
7 мм
0,10
0,02
0,12
5,63
ошибка
15
MULTICOLOR
(HIBOTAN)
старые
ткани
0,10
0,16
0,26
0,57
0,54
16
PINK
бутон
0,07
0,03
0,10
2,41
2,17
17
RED
детка
7 мм
0,04
0,19
0,23
0,15
0,63
В
среднем по группе
0,47
0,25
0,73
2,07
1,98
Стандартное
отклонение
0,43
0,17
0,57
1,65
1,22
Наиболее удивительно, что такие АЦФ как MULTICOLOR и ВLACK содержат заметно больше хлорофилла, чем растущий на собственных корнях исходный вид Gymnocalycium mihanovichii var friedrichii (0,48-1,54 мг/г против 0,45 мг/г). Это свидетельствует о том, что такие АЦФ ни в коем случае нельзя называть "бесхлорофильными" и даже "малохлорофильными". Вероятно, они не так уж и нуждаются в прививке. Их вполне можно выращивать на собственных корнях. Все прочие АЦФ содержат значительно меньше хлорофилла, чем Gymnocalycium mihanovichi var. friedrichii, от 0,35 у эриокактуса до 0,04 у АЦФ "RED". Пожалуй, только эту темно-красную АЦФ можно с оговорками признать "истинно бесхлорофильной".
Аналогичные результаты получаются и по сумме пигментов: MULTICOLOR и ВLACK чаще имеют показатели 1-2 мг/г против 0,82 у исходного вида и 0,1-0,4 мг/г у большинства остальных АЦФ.
Большие различия наблюдаются по всем параметрам и у разных частей одного и того же растения, которые генетически являются однородными. Это хорошо заметно на примере АЦФ MULTICOLOR, у которой образуются детки разного цвета - от почти чисто черных, до почти чисто оранжево-красных. Соответственно, у первых пигментов, особенно хлорофиллов почти в 2 (!) раза больше чем у вторых. Далее, в детках содержание пигментов выше, чем в молодых тканях (приросте текущего года) материнского растения, а в этих последних, соответственно, в 5-6 раз (!) выше, чем в нижележащих старых тканях 4-5-летнего возраста. Многократно высказанная в литературе мысль об ускоренном старении АЦФ здесь находит прямое подтверждение: с увеличением возраста ткани кактуса происходит быстрое (чуть ли не линейное) снижение содержания фотосинтетических пигментов, что, однако, требует дополнительного изучения.
В бутонах АЦФ MULTICOLOR содержится больше хлорофилла, чем в молодых и, тем более, старых соматических тканях, в то время как содержание каротиноидов от такового в молодых тканях не отличается, хотя в 2 раза превышает его в старых тканях. В бутонах АЦФ BLACK, напротив, хлорофилла и каротиноидов меньше, чем в молодых тканях кактуса. В бутонах и детках формы PINK содержание хлорофиллов и каротиноидов идентично. В общем же явной зависимости концентрации пигментов в генеративных органах от содержания их в соматической ткани кактуса пока не обнаружено, хотя возможно она и существует.
Соотношение "хлорофилл : каротиноиды" у АЦФ очень изменчиво (0,15-6,08). На примере той же формы MULTICOLOR можно показать, что оно может быть даже больше, чем у нормально фотосинтезирующего исходного вида G. mihanovichii var. friedrichii, хотя в старых тканях MULTICOLOR и этот индекс почти в два раза снижается по сравнению с детками и молодыми тканями и в 6 раз - по сравнению с бутонами.
Соотношение хлорофиллов a и b также в высшей степени изменчиво (0,53-4,24), причем оно столь же сильно варьирует и у разных тканей одной АЦФ MULTICOLOR (0,54-4,24).Столь сильные различия в тканях одного кактуса свидетельствует о значительной связи этого показателя с изменением условий окружающей среды. Удивительно, что у исходного вида, "нормального" гимнокалициума, это соотношение составляет 0,53, в то время как у самых тенелюбивых сциофитов этот показатель должен быть никак не меньше 2,3.
ВЫВОДЫ
Цветные формы кактусов имеют аномально низкое содержание хлорофиллов. У суккулентов семейства толстянковых содержание хлорофиллов ненамного больше, в то время как у обычных лиственных растений их количество, по меньшей мере, на порядок выше.
Содержание каротиноидов у цветных форм, кактусов-подвоев и толстянковых практически не отличается. Это однозначно свидетельствует о том, что окраска цветных форм кактусов вообще, и ее интенсивность в частности, не связана с повышенным содержанием каротиноидов. У лиственных растений содержание каротиноидов и сумма пигментов также на порядок выше.
Отношение "хлорофиллы : каротиноиды" у цветных форм оказалось наименьшим среди изученных групп, и вообще исключительно низким. Различия между подвоями, толстянковыми и лиственными растениями по этому показателю незначительны. Кактусы-подвои приближаются скорее к сциофитам, а толстянковые занимают лишь промежуточное положение между гелиофитами и сциофитами.
Отношение "хлорофилл а : хлорофилл b", у цветных форм по сравнению с подвоями и лиственными растениями существенно меньше, хотя и незначительно отличается по сравнению с толстянковыми.
Выявлены существенные различия по содержанию пигментов и их индексам у разных частей одного и того же растения, что четко показано на примере АЦФ MULTICOLOR. В детках содержание пигментов выше, чем в молодых тканях материнского растения, а в них, соответственно, в 5-6 раз выше, чем в нижележащих старых тканях 4-5-летнего возраста. Эти данные прямо подтверждают мысль об ускоренном старении цветных форм, при котором происходит быстрое снижение содержания фотосинтетических пигментов.
Большинство цветных форм некорректно называть "бесхлорофильными", поскольку хлорофиллы в них содержатся, хотя бы и в незначительном количестве, а иногда и в большем, чем у исходного корнесобственного растения Gymnocalycium mihanovichii var. friedrichii.
ЛИТЕРАТУРА
Горышина Т.К. Экология растений: Учебное пособие. М.: Высшая школа, 1979. с.9-46.
Калам Ю., Орав Т. Хлорофильная мутация. Таллин, 1974. 58 с.
Калишев В.Б. Цветные формы кактусов. Сборник статей. Челябинск, 1999. 40 с.
Либберт Э. Физиология растений. М.: Мир, 1976. 456 с.
Насыров Ю.С. Фотосинтез и генетика хлоропласта. М.: Наука, 1975. 143 с.
Овчинников Н.Н., Шиханова Н.М. Фотосинтез. Пособие для учителей. М.: Просвещение, 1972. с. 71-91.
Walles B. In: Structure and function of chloroplasts/ Berlin; Heidelberg; N.Y., 1971. p.51.
Cultivar e-magazin: Copyright (c) by Valery Kalishev, Chelyabinsk, Russia, since 2000. Design and hosting by Peter Lapshin, since 2002. Contacts: Peter Lapshin