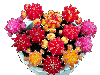Crossing Experiments Within the Genus Ariocarpus. Scheidweiler
T. Neudecker (photos by the author)
This article is reproduced form B.C.S.S.Journal Volume 3 (1) 14-17 (1985) with the kind permission of the Editor. The link to sourse of this article here.
The genus Ariocarpus is certainly one of the most interesting of the cactus family. Due to the strange appearance of its species it has gained much attention both by growers and by botanists ever since the description of Ariocarpus retusus , the type species of the genus, by Scheidweiler in 1838. Until Anderson's revision in 1965 the six generally accepted species (and a few varieties or forms) of Ariocarpus were usually split into three different genera: (a) Ariocarpus itself with the species retusus, trigonus and scapharostrus , (b) Roseocactus , a concept of Berger (1925), including the species fissuratus and kotschoubeyanus which are characterized by a central woolly groove on their flattened triangular tubercles, and (c) the monotypic genus Neogomesia , erected by Casta?eda in 1941 for his discovery of the species agavoides on a hill near the Mexican town of Tula, Tamaulipas. It was Anderson's fundamental work that finally led to the unifying concept, now generally accepted, of a genus Ariocarpus comprising all the species mentioned above and treating Roseocactus and Neogomesia as mere subgenera at the most.
It was this concept of Anderson that finally induced me to start this survey by systematically doing crossing experiments within the genus Ariocarpus , not only out of pure curiosity as to what can be achieved by intentionally hybridizing such interesting plants, but also to get information regarding genetic compatibility of Ariocarpus species of the same as well as different subgenera and possibly even of Ariocarpus and related genera such as Encephalocarpus, Obregonia, Strombocactus and Leuchtenbergia .
The first question to be answered was this: Are there genetic barriers between the species of the different subgenera of the genus? If not, this could be regarded as a strong argument in favour of Anderson's concept. Secondly, it seemed most interesting to watch how specific characteristics of certain species turn up or are suppressed in hybrid plants. It must be mentioned here that Anderson himself conducted crossing experiments to support his unifying concept and reported on the production of viable seed in several cases. However, he obviously did not raise hybrid seedlings in order to compare the appearance of older plants with that of their respective parents.
Considering the possibility of pseudo-successful hybridizing, e.g. due to apomixis (stimulation of female reproductive cells to develop embryos without fertilization) - a phenomenon known to occur in several plant families - the production of viable seed by a cross-pollinated mother plant does not necessarily prove the success of a crossing experiment unless the F1 generation really shows hybrid characteristics. Such characteristics, however, cannot be observed in seedling plants of Ariocarpus . This was another important reason for actually raising Ariocarpus plants resulting from crossing experiments
At first sight, such a project seems to be extremely time-consuming due to the exceedingly slow growth of Ariocarpus from seed. Specimens which are grown on their own roots from the beginning cannot be expected to bloom before they are ten years old or more, even under optimal conditions. This difficulty, however, can be circumvented elegantly by grafting Ariocarpus seedlings onto Pereskiopsis . Such a procedure usually leads to an enormous growth rate of the graft which can reach a diameter of 3 cm or more and regrafted on Eriocereus (Harrisia) j usbertii may bloom for the first time at the age of two to three years. Under such conditions there are good chances not only of getting primary hybrids (F1 generation) between different species, but also of obtaining re-crossings and eventually hybrids with three, four or even more parent species within a reasonable time.
The first 3 parent hybrid seedlings of Ariocarpus were raised in 1984. They will be the topic of another report. This survey will be dealing only with primary hybrids clearly showing characteristics intermediate between those of their two different parent species. Some of these hybrids have recently produced their first remarkable flowers.
Figure 1 shows all the successful crossings which have been performed up to now (1984) and have resulted in hybrid specimens growing in my collection. For reasons of clarity, varieties and mere forms of the six species of Ariocarpus have not been differentiated. This means that A. fissuratus and its variety Iloydii are treated as one crossing parent, as well as A. retusus and its variety (forma?) furfuraceus . The same is true for A. kotschoubeyanus and its forms macdowellii and 'elephantidens ". Only in the case of A. kotschoubeyanus f. albiflora an exception from the rule seemed to be justified in so far as its white (instead of wine-red) flower obviously influences the color of the hybrid flower in a very different way. Thus the crossing experiments have been performed with six species and one forma:
A. agavoides (=agav.), A. fissuratus (including Iloydii ) (=fiss), A. kotschoubeyanus (including macdowellii and 'elephantidens' ) (=kots.), A. retusus (including furfuraceus ) (=ret.), A. scapharostrus (=scaph.), A. trigonus (=trig.) and A. kotschoubeyanus f. afbiflora (=kots. a.).
| FEMALE |
MALE |
| agav. |
fiss. |
kots. |
kots. a. |
ret. |
scaph. |
trig. |
| agav. |
|
|
X |
X |
X |
X |
X |
| fiss. |
X |
|
|
X |
X |
X |
X |
| kots. |
X |
X |
|
X |
X |
X |
X |
| kots. a. |
|
|
|
|
X |
X |
X |
| ret. |
X |
X |
X |
|
|
X |
X |
| scaph. |
X |
|
|
|
|
|
|
| trig. |
X |
X |
X |
X |
|
X |
|
Fig 1: Successful crossings and raisings of hybrid plants (blue/X areas) up to1984
A closer look at Figure 1 reveals that up to now at least one out of each two (reciprocal) possible hybrids could be achieved; in some cases both reciprocal crossings have already been performed. For example: hybrids combining the genetic material of both A. agavoides and A. retusus could be obtained both in the form of ret . x agav . and agav . x ret . (female parent first). As there is a strange tendency in Ariocarpus hybrids to display morphologically more characteristics of the male (i.e. pollen-donating) than of the female parent - a fact which has also been observed by Lux and Stanik (1983), reciprocal hybrids of the same two parents often differ remarkably in their appearance.
Another point has to be considered: according to Figure 1 A. scapharostrus , for example, served only once as the female parent, but was male parent in all other possible crossings. Of course this fact must not be interpreted as an indication of some strange genetic incompatibility, it merely reflects the difficulty of getting enough plants of this rare species into bloom. Once in flower, one such flower can easily serve as the pollen donator (male parent) for many different crossings while, for reasons of clarity, serving only one as the female parent. So it seems only a matter of time before these and the other gaps in the planned scheme can be filled.
Despite its incompleteness Figure 1 gives sufficient information for this important statement: obviously all the species of Ariocarpus are genetically compatible, no matter what subgenus they belong to. This is a strong experimental evidence for a very close genetical and evolutionary relationship between all Ariocarpus species, an observation which gains importance in the light of the fact that so far-despite many trials-all crossing experiments of Ariocarpus with Encephalocarpus, Obregonia, Strombocactus and Leuchtenbergia have failed to produce viable seed.
The next question to be answered was this: can crossing experiments further elucidate the degree of interspecies relationship within the genus Ariocarpus ? As has often been observed by Lux and Stanik, Ariocarpus crossings very often yield seedlings with obvious defects in their chlorophyll synthesizing system. In some cases practically all the plantlets show chlorophyll deficiency, i.e. they are either white or red in varying intensities. Sometimes even attractive `variegated' specimens can be observed. Such defective seedlings may survive by early grafting, although the 'taking' rate of such grafts is considerably lower than with normal green plants. Furthermore, it is not unusual to see initially white or red grafts finally turning green, a phenomenon which might be the result of crucial gene products not synthesized in the graft itself, but being provided by the green stock.
The crucial question is whether parameters such as the number of seeds produced per capsule or the percentage of chlorophyll-deficient seedlings resulting from a crossing experiment can be considered a reliable measure of a more or less close genetic relationship between the two parent species. Seed production is likely to be influenced by too many other parameters such as temperature during pollination, age of pollen, physiology of the stigma etc. to serve as a clear indicator of genetic relationship. The biochemical parameter of chlorophyll synthesis in seedlings produced seems much more adequate as such an indicator. However, experience gained so far in this type of experiment does not seem to support a simple correlation between the percentage of chlorophyll-deficient specimens in a population of hybrid seedlings and the genetic relations of the parent species.
This can be illustrated by the following observation. The crossing ret . x fiss , yielded many viable seeds per capsule, whereas the reciprocal crossing fiss . x ret . produced only a few seeds: in both cases all the seedlings of this cross between species belonging to two different subgenera were green. On the other hand, the crossing kotsch . x fiss . (both parents belonging to the same subgenus and obviously rather closely related) produced a lot of viable seed, but none of the seedlings appeared to be fully green. Another example: trig . x fiss . is a crossing easily obtained, yielding a large percentage of normal green colored seedlings; trig . x scaph ., however, produced only a few seeds of poor viability and all the seedlings appeared to be chlorophyll-deficient.
These examples clearly demonstrate that the efficiency of crossing experiments within the genus Ariocarpus is- at least partly determined by factors other than genetic relationship alone. This situation might be further complicated by the possibility-preliminary experimental evidence has been obtained already-that even within one species different clones of different compatibilities in cross fertilization may exist. This question has to be solved in further experiments.
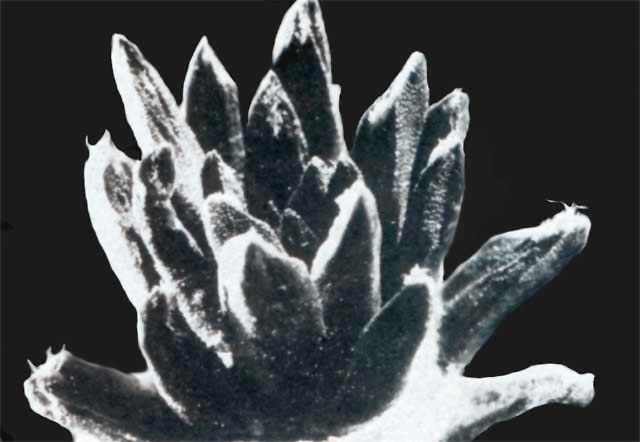
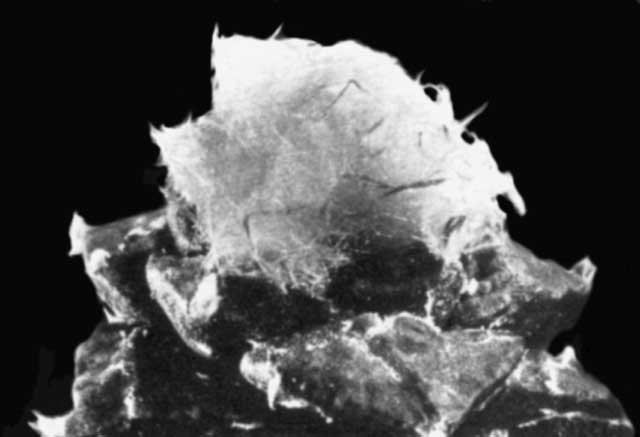
The pictures accompanying this article are intended to give a general impression of the astonishing variability of new forms which can be obtained by hybridizing Ariocarpus species. More detailed observations on certain crossing groups and the influence of certain parent species will be discussed in forthcoming articles.
References
ANDERSON, E. F. (1961) A taxonomic revision of Ariocarpus, Pelecyphora and Obregonia (Family Cactaceae). Dissertation, Claremont Graduate School.
ANDERSON, E. F. (1965) A revision of Ariocarpus . Cact. Succ. J. Amer. 37: 39-49.
LUX, A. and STANIK R. (1983) Hybridisation in der Gattung Ariocarpus Scheidweller. Kak. u.a. Sukk. 34: 267-271.
Illustrations: (Will open in a new window)
In the captions to all illustrations the female parent is given first
The quality of some of the illustrations accompanying this article, does not meet with today's more exacting standards. This is due to shortcomings in the reproduction of the original article