Origin, Development and Propagation of Chimeras
R. Daniel Lineberger, Professor of Horticulture
Department of Horticultural Sciences, Texas A&M University
INTRODUCTION
A plant is said to be a chimera when cells of more than one genotype (genetic makeup) are found growing adjacent in the tissues of that plant. Variegated plants are perhaps the most common types of chimeras, and certainly the most convenient example to use in presenting the basic concept. The cells in a variegated leaf all originated in the apical meristem of the shoot, but some cells are characterized by the inability to synthesize chlorophyll. These appear white rather than green even though they are components of the same tissue system. Many important selections of foliage, floricultural, and landscape plants are chimeras. Cornus alba 'Argenteo Marginata', Vinca minor 'Variegata', Ajuga reptans 'Burgundy Glow', and many selections of Hosta, Diffenbachia, Peperomia, Chlorophytum, and Saintpaulia are but a few of the chimeras which have variegated foliage.
The intent of this article is to explore the origins and development of chimeral plants, to introduce precautions which must be followed in the propagation of chimeras, and to discuss horticulturally significant chimeras other than variegated foliage types.
THE CONCEPT OF APICAL ORGANIZATION
No discussion of the origin of chimeras would be complete without a review of the organization of the shoot apex. The pattern of cell division, frequency of cell division, and layered organization of the cells in the apex interact in determining the type of chimera which is produced and the stability of the pattern which results.
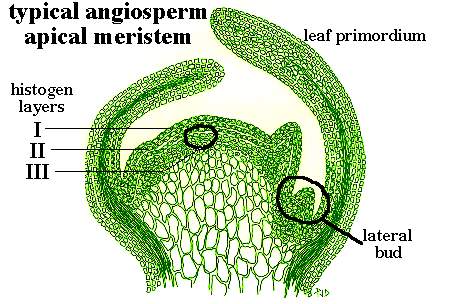
The apical meristem of a shoot is the location where most of the cells which produce the plant body are formed. Cell division occurs at a very rapid rate in an actively growing shoot and these cells in turn elongate or expand resulting in growth in length of the shoot. Woody plants and certain herbaceous plants contain secondary meristems which result in increase in growth or caliper, but this secondary growth will not be considered here. On photo 1 is a sketch of a longitudinal (or lengthwise) section through the apical meristem of a typical woody or herbaceous dicot shoot. Leaf primordia arise on the sides of the apical dome, and lateral buds develop in the axils of these young leaves.
The apex is organized into a layered region (the tunica) and a region where layering is not evident (the corpus). The controlled pattern of cell divisions in the tunica results in the maintenance of discrete layers, with the number of layers varying somewhat among the different species. It is significant to note that the layers retain their organization into the region where the leaves and lateral buds are developed.
The derivatives (or progeny, if you will) of the outermost layer (L.I) give rise to the epidermis (Photo 1). The epidermal layer is continuous as an outer covering over all tissues of the leaf, stem, flower petals, etc. Derivatives of layer II (L.II) give rise to several layers within the stem and a large proportion of the cells in the leaf blade. Derivatives of layer III (L.III) give rise to most of the internal tissue of the stem and a number of cells around the veins within the leaf. The significance of the cell layers and their resulting progeny will be discussed in more detail below.
THE ORIGIN OF CHIMERAS
Chimeras arise when a cell undergoes mutation. This mutation may be spontaneous or it may be induced by irradiation or treatment with chemical mutagens. If the cell which mutates is located near the crest of the apical dome, then all other cells which are produced by division from it will also be the mutated type. The result will be cells of different genotypes growing adjacent in a plant tissue, the definition of a chimera.
If the location of the cell at the time of mutation is in a region where little further cell division will occur, then the likelihood of detecting this mutation by visual inspection of the whole plant is low. Furthermore, if the mutation results in a genotype which is not very different morphologically from the rest of the plant, then the likelihood of identifying the plant as a chimera is also low. A mutation which results in colorless rather than green cells (variegation) is easily detectable, whereas a mutation which results in greater sugar accumulation in the cells would not be observed.
Periclinal, Mericlinal, and Sectorial Chimeras
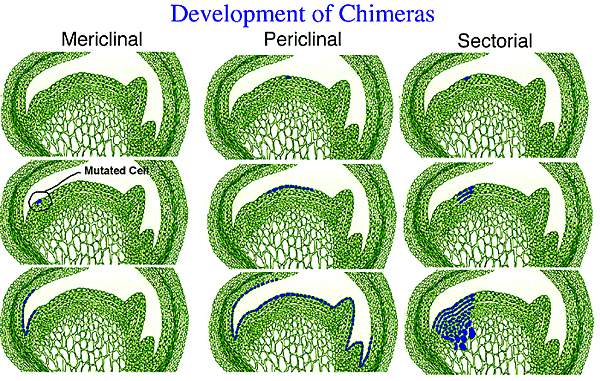
Chimeral plants can be categorized on the basis of the location and relative proportion of mutated to nonmutated cells in the apical meristem. Periclinal chimeras (Photo 2) are the most important category since they are relatively stable and can be vegetatively propagated. A mutation produces a periclinal chimera if the affected cell is positioned near the apical dome so that the cells produced by subsequent divisions form an entire layer of the mutated type. The resulting meristem contains one layer which is genetically different from the remainder of the meristem. If, for example, the mutation occurs in L.I, then the epidermal layer of the shoot which is produced after the mutation is the new genetic type.
A classical example of an L.I periclinal chimera is the thornless blackberry. The epidermal layer of this type produces no "thorns" (the modified epidermal cells are correctly called "prickles"). The thornless epidermis covers a stem whose cells contain the information for the thorny genotype. This can be demonstrated by taking root cuttings. The adventitious shoots which differentiate on the root cuttings are not chimeral and therefore revert to the thorny genotype.
Mericlinal chimeras (Photo 2) are produced when the derivatives of the mutated cell do not entirely cover the apical dome. A mutated cell layer may be maintained on only one portion of the meristem giving rise to chimeral shoots or leaves which develop in that portion while those that differentiate on all other portions of the meristem are normal, nonchimeral shoots. Many mericlinal chimeras involve such a limited number of cells that only a small portion of one leaf may be affected. As was the case with periclinal chimeras, mericlinal chimeras are generally restricted to one cell layer.
Sectorial chimeras (Photo 2) result from mutations which affect sections of the apical meristem, the altered genotype extending through all the cell layers. This chimeral type is unstable and can give rise to shoots and leaves which are not chimeras. Both normal types and mutated types can be produced, depending upon the point on the apex from which the shoots differentiate.
RECOGNITION OF CHIMERAL PATTERNS
Careful observation of the pattern of coloration in variegated dicot leaves allows one to partially deduce the nature of the chimeral layers in the shoot apex. A complete characterization of the nature of all three layers cannot be made macroscopically since most of the cells of the epidermal layer of dicot leaves do not produce chloroplasts even if the genetic information for chloroplast development is present. However, the guard cells of the stomatal apparatus do contain chloroplasts in a genetically "green" L.I, but these must be observed microscopically.
The terminology used to describe the nature of the apical cell layers is to use upper case letters in sequence to denote the genetic composition of the cells contained therein. For example, the correct designation for a periclinal chimera with a genetically green L.I, white L.II, and green L.III is G-W-G.
In a dicot leaf, the margins of the leaf are derived from L.II. The extent of the leaf margin contributed by L.II is variable from species to species, and to some extent variable among the leaves on an individual stem. The leaf pictured in Figure 2A was produced by a meristem which was G-G-G (nonchimeral, not variegated), while the leaf in Figure 2B was produced by a G-W-G variegated sport.
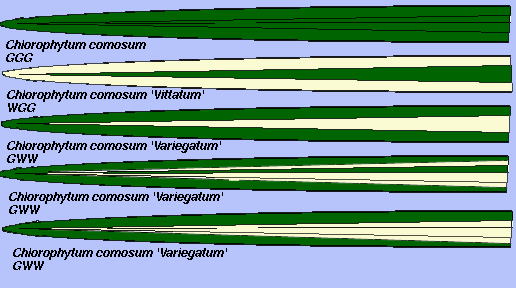
In monocotyledenous species, L.I does contribute to the development of the leaf margin and the majority of the central portion of the leaf develops from L.II. Unlike dicot leaves, L.III contributes only a small proportion of the cells of the leaf blade. Figures 3 A-E represent leaf variegation patterns of Chlorophytum comosum. Figures 3A (top) is from a G-G-G (nonchimeral, not variegated) plant, Figure 3B (second from top) is from a W-G-G plant, and Figure 3C (middle) is from a G-W-W plant. The leaf to leaf variability in the contribution of L.I and II to the proportion of cells in the leaves of an individual plant is demonstrated in Figures 3D and 3E (bottom two leaves). Both leaves were taken from the same plant, but the marginal green area contributed by L.I to the leaf in Figure 3D was much greater. Such observations indicate that the development of leaf patterns, and therefore the contributions of the different apical cell layers to leaf development, are not rigidly controlled.
MANIFESTATIONS OF CHIMERAS OTHER THAN VARIEGATION
While variegation represents a very visible manifestation of chimeras, it should be apparent from the previous discussion that in theory a plant could be a chimera for virtually any trait. Other frequent chimeras involve loss of epidermal appendages (thornlessness in blackberries, "fuzzless" peaches), alteration in bract color in poinsettia, and various petal or flower color patterns in carnation and chrysanthemum.
Chimeras for ploidy level have been extensively studied in fruit crops. Tetraploidy (doubling of the normal chromosome number) results in fruit which is much larger than normal when the trait is located in L.II and L.III. This phenomenon has been studied in apples and grapes.
PROPAGATION OF CHIMERAL PLANTS
Mericlinal and sectorial chimeras are by their very nature unstable and the likelihood of propagating plants with the same morphological pattern from these types is low. Periclinal chimeras are very stable and in some cases (as in variegated Chlorophytum) the chimeral plant is the most frequently available form in commerce.
Most commonly used techniques for plant propagation result in the formation of true-to-type periclinal chimeras. This fidelity of propagation accrues from the fact that the arrangement of the cell layers in the apical meristem of lateral buds is identical to the arrangement of the meristem from which they were formed. Thus layers I, II, and III contribute cells to the differentiating lateral meristems. Rooted cuttings, budding, grafting, divisions, and tip layering are all high fidelity propagation practices. Important departures from the rule are found in cases where the propagules differentiate from adventitious shoots. A case discussed previously which illustrates this point is the thornless blackberry. Adventitious roots which form on a blackberry stem cutting or tip layer originate in the subepidermal tissues of the stem (L.II and L.III). If root cuttings are then taken from these plants, the adventitious shoots do not contain the layer with the thornless genotype (L.I) and the propagules are of the thorny type.
The "pinwheel flowering" African violets (Photo 4) similarly cannot be propagated in a true-to-type fashion from leaf cuttings. Shoots which originate on the leaf cuttings are either a single color or irregularly mottled bicolors - (Photo 5) without the characteristic flowering pattern. High fidelity propagation of the pinwheel flowering African violets is currently achieved by separating "suckers" which originate from lateral buds. This illustrates that the characteristic flowering pattern of the cell layers in the apical meristem.
CHIMERAL SEPARATION IN TISSUE CULTURE
Plant tissue culture allows the propagation of plants from very small numbers of cells within a tissue system or in the extreme case, from single isolated cells (protoplast culture). When applied to chimeral plants, tissue culture technology offers an unprecedented tool for "dissecting" or separating chimeras for morphological study of the component genotypes. In like manner, the fact that chimeras may separate due to adventitious shoot formation with the concomitant disruption of the orderly arrangement of the apical cell layers poses serious problems in using this technique for rapid clonal propagation.
Experience in our laboratory has shown that Ajuga reptans 'Burgundy Glow' proliferates in vitro from both axillary buds and adventitious shoots - (Photo 6). Those plantlets which arise from axillary buds are of the normal 'Burgundy Glow' phenotype. However, off-type plantlets are also produced. These are of two types. The most frequent type is the ТbronzeУ or uniformly deep green foliage type. The other type is completely devoid of chlorophyll, appearing pink in culture, but failing to root and grow due to the inability to carry on photosynthesis. Most proliferation occurs via axillary bud development since nearly 70% of the plantlets produced are true-to-type 'Burgundy Glow' - (Photo 7).
SUMMARY
Stable periclinal chimeras have resulted in plant sports with unique ornamental features. The ability to propagate these sports in a true-to-type fashion depends upon the use of a propagation technique which utilizes lateral buds. The layered organization of the apical meristem which is characteristic for a periclinal chimera is maintained in lateral meristems, but is usually lost during the differentiation of adventitious shoots. Plant tissue culture methodology enables the separation of chimeral genotypes in some species, facilitating the study of the separated component genotypes. This advantage of tissue culture may be a liability in systems which proliferate in vitro through adventitious shoot formation, since the propagation of off-type plants may occur.
SUGGESTIONS FOR FURTHER READING
- Dermen, H. 1955. A 2-4-2 chimera of McIntosh apple. J. Wash. Acad. of Sci. 45:324-327.
- Dermen, H. 1960. Nature of Plant Sports. The Horticultural Magazine, July 1960, pp. 123-173.
- Esau, K. 1965. Plant Anatomy. Second Edition, John Wiley & Sons, Inc., New York, pp. 89-112.
- Stewart, R.N. and H. Dermen. 1979. Ontogeny in monocotyledons as revealed by studies of the developmental anatomy of periclinal chloroplast chimeras. Amer. J. Bot. 66, pp. 47-58.